This article aims at reconstructing history of R1a1 ancient migrations between 20,000 and 3500
years before present (ybp). Four thousand four hundred sixty (4460) haplotypes of haplogroup R1a1
were considered in terms of base (ancestral) haplotypes of R1a1 populations and timespans to their
common ancestors in the regions from South Siberia and northern/northwestern China in the east to
the Hindustan and further west across Iranian Plateau, Anatolia, Asia Minor and to the Balkans in
Europe, including on this way Central Asia, South India, Nepal, Oman, the Middle East, Comoros
Islands, Egypt, etc. This study provides a support to the theory that haplogroup R1a arose in
Central Asia, apparently in South Siberia and/or neighboring regions, around 20,000 ybp.
(Apparently, the most ancient source of R1a1 haplotypes is provided by the people now living in
northern China, at approximately 20,000 ybp). Not later
than 12,000 ybp bearers of R1a1 already were in the Hindustan, then went across Anatolia and the
rest of Asia Minor apparently between 10,000 and 9000 ybp, and around 9000 - 8000 ybp they arrived
to the Balkans and spread over Europe east to the British Isles. On this migration way or before it
bearers of R1a1 (or the parent, upstream haplogroups) have developed Proto Indo-European language,
and carried it along during their journey to Europe. The earliest signs of the language on passing
of bearers of R1a1 through Anatolia were picked by the linguists, and dated by 9400 - 9600 - 10,100
ybp, which fairly coincides with the data of DNA genealogy, described in this work. At the same time
as bearers of the brother haplogroup R1b1a2 began to populate Europe after 4800 ybp, haplogroup R1a1
moved to the East European Plain around 4800 - 4600 ybp. From there R1a1 migrated (or moved as
military expeditions) to the south (Anatolia, Mitanni and the Arabian Peninsula), east (South Ural
and then North India), and south-east (the Iranian Plateau) as the historic legendary Aryans.
Haplotypes of their direct descendants are strikingly similar up to 67 markers with contemporary
ethnic Russians of haplogroup R1a1. Dates of those Aryan movements from the East European Plain in
said directions are also strikingly similar, between 4200 and 3600 ybp.
The first writing system, the original Mesopotamian (believed to be the world's oldest) has emerged
circa 3600 BC (5600 ybp), it belonged to an agglutinative language, and any notion of a flexive
Proto Indo-European language, or for that matter any language, known from the 9400 - 9600 - 10,100
ybp hangs in fine etheric vacuum, next to the theories of unintelligent and intelligent creation.
The first written materials ascribed, with reservations, to an Indo-European language date to the 17th
c. BC (3700 ybp, Anatolia), any references before that belong to the I.Asimov's-type compositions.
Very few linguists would bet a half-dollar on anything older than 5700 BC. In fact, the Proto
Indo-European language notion is the creationist model of a single-origin family tree, one of the
few theories espoused by the linguists; without a Wave model, it even does not hold water.
However, the credulous attitude of the authors is irrelevant to the authors' findings on the R1a
march in time and space.
|
Introduction
This study focuses on the origin of Indo-Europeans and the Aryans who entered India (the
Hindustan), Iran (Iranian plateau), and Anatolia (Mesopotamia) approximately 3500 years ago.
The research findings, described in this study, demystify the origin of the Aryans. For nearly
two centuries, the "Aryan problem" (essentially — Who were the Aryans? Where did they come from?
Where did they disappear? Were they a particular human race, different from others?) has posed many
challenges, often controversial and conflicted, for researchers, archeologists and linguists;
however, this study opens new ground for our consideration and is based on the data provided by DNA
genealogical test results.
The methodology of DNA genealogy, including considerations of extended 67 marker haplotypes, is
described in detail in the preceding paper in this journal (Rozhanskii & Klyosov, 2011) and in
Materials and Methods section of this article. The 67 marker haplotypes have been introduced to the
scientific domain and personal usage several years ago, and available databases containing tens of
thousands of 67 marker haplotypes are listed in (Rozhanskii & Klyosov, 2011) and in this paper
(Appendix).
First, the following two 67 marker haplotypes of haplogroup R1a1 are presented, belonging to the
two authors of this paper:
13 24 16 11 11 15 12 12 10 13 11 30 — 16 9 10 11 11 24
14 20 34 15 15 16 16 — 11 11 19 23 15 16 17 21 36 41 12 11 — 11 9 17 17 8 11 10 8 10 10 12 22 22 15
10 12 12 13 8 15 23 21 12 13 11 13 11 11 12 13
13 25 15 10 11 14 12 12 12 13 11 30 – 16 9 10 11 11 23
14 20 34 12 12 15 15 — 10 11 19 23 17 16 18 18 34 38 14 11 — 11 8 17 17 8 11 10 8 12 10 12 21 22 15
10 12 12 13 8 13 25 21 13 12 12 13 11 11 12 13
Next, the following are two Indian R1a1 haplotypes, taken arbitrarily from the "Indian FTDNA
Project" (the references to Projects are at the end of this paper):
13 24 16 11 11 14 12 12 10 13 11 31 — 16
9 10 11 11 24 14 20 33 12 15 15 16 — 10 12 19 23 15 17 18 18 35 41 15
11 — 11 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 13 23 21 12
12 11 13 10 11 12 12
13 23 16 11 12
15 12 12 10 13 11 30 — 16 9 10 11 11 24 14 20 30 12 16 16 16 — 11
12 19 23 15 16 18 21 35 39 12 11 — 11 8 17 17 8 12 10 8 11
10 12 22 22 16 10 12 12 13 8 14 24 22 13 13 11 13 11 11 12 12
1
Both Indian haplotypes contain 24 and 21 mutations with the first haplotype (mutations are shown
in bold, and rules of their counting are explained in the paper cited above), and 28 and 36
mutations with the second one. This produces on average 27.25 ± 6.60 pairwise cross-mutations
between all four haplotypes; that is, 27.25/.12 = 227 → 292 ± 55 conditional generations (25 years
each) = 7300 ± 1400 years between two haplotypes on average, or 3650 ± 700 years to a common
ancestor of all the four haplotypes (.12 here is the mutation rate constant for 67 marker
haplotypes, see the preceding paper cited above). According to all historical accounts, the Aryans
arrived in India in the middle of the 2nd millennium BC, which is approximately 3500 years ago.
This simplified calculation is based on these four haplotypes that belong to different subclades
of R1a1 haplogroup (Z280, M458 and L342.2). However, considering each of the four haplotypes, the
first two are from the current Russian-Ukrainian (linguistically
Indo-European) group, and the second two are from the Indian (linguistically
Indo-European) group. Both are similar and belong to the same R1a1 haplogroup. Currently, up to 72%
of the upper castes in India belong to bearers of the same R1a1 haplogroup (Sharma et al., 2009).
This simplified calculation is given here for illustrative purposes, though four 67 marker
haplotypes contain as many as 268 markers, which is quite statistically informative in a first
approximation. A much more detailed analysis of Indian and ethnic Russian extended series of R1a1
haplotypes is given in (Klyosov, 2009b, 2011b), and principally the same results were obtained with
respect to patterns of mutations in haplotypes, migration routes, and their chronology.
This brings closure to the question of the Aryans’ DNA-related origin and who entered India
during the middle of the 2nd millennium CE. They belonged to the R1a1 haplogroup, which is the
prevalent one in the present-day Eastern Europe (Russia, Poland, Ukraine, Belarus, in the first one
up to 62% of total male population, in the latter three up to 55% of total male population [Klyosov,
2009b, 2011b and references therein]).
| The reverse projection of the population genetics' results onto the present-day Eastern
Europe (Russia, Poland, Ukraine, Belarus) equates the modern location of the Slavic-speaking people
with the modern distribution of the Y-DNA marker. In the historical terms, such equating comes in
conflict with the distribution of the people belonging to different linguistic trunks, as is
illustrated on the maps shown below: according to the modern interpretation of the Ptolemy's
distribution of the tribes, the recorded population of the Eastern Europe at the verge of the eras
was predominantly Türkic-Scythian-Sarmatian in the southern steppes and Fennic in the forest areas,
the Scythians were nomadic horse pastoral tribes with little in common with the Aryan (Indo-Iranian)
sedentary agricultural tribes. A look at the map shows that either the population was nearly
completely displaced, together with their genetic signatures, or assimilated to the disappearance of
free pasture ranges, new dominant language, and new culture. In either case, the equation of the
current Eastern European linguistic siblings of the Indian Brahmans with the genetic siblings of the
Indian Brahmans runs into a conflicting collision. What used to be a chain of migratory events from
20,000 ybp according to the DNA-genealogy becomes an annoyingly static picture after the R1a reached
the promised land in the E. Europe ~6000 ybp. That also conflicts with the Slavic annals, where
Slavs migrated to the E. Europe from the Danube area. The ancestral Slavs in the Danube basin area are not noted for their genetic kinship with their
descendent Eastern European Slavs, they are of different ancestry: predominant haplogroup is I
at Bosnians (42%), Croats (mainland) (38%), Herzegovinians (63%), Ethnic Macedonians (34%),
Macedonians (Skopje) (23%), Serbs (36%), Serbs (Bosnia) (40.7%), Slovenians (30.7%), Slovenian
(38%);
E1b1b at Bulgarians (20%);
R1b at Czechs and Slovaks (35%).
These genetic differences can be attributed to different historical causes in the last 1500 years
for each Slavic group with predominant haplogroups I, E1b1b, and R1b, but then
the same last 1500 years logics should apply to those Slavs bearing the haplogroup R1a.
One of the historical scenarios for the explosive spread of R1a in the Eastern Europe may be
Droit du seigneur (Right of first night), that survived in Russia well into the 19th century.
The relatively small ruling elite, and possibly all members of the demographically small dominating
ethnic group, were seeding into posterity their Y-DNA marker that originated some 4,500 ybp,
speedily and efficiently replacing the original Slavic markers. For centuries, the Rus and Russian
standing army primarily consisted of Slavicized elite inherited from the Bulgar and Khazar states,
and of the allied Türkic tribes (Black Clobuks, Oguzes, Baryns, etc.), "boyar offsprings", and
"Tatars" whose ethnicity automatically classed them as service nobility, and the pay for the service
was by land allotments with permanently bonded occupants. The heavy dose of non-Slavic nobility able
to seed their seeds is attested by the predominance of the Türkic-derived elite family names:
Akchurins, Arginskys (from tribe Argyn, a part of annalistic Basmyls), Barynskys (dynastic tribe of
Savirs), Bichurins, Karamzins, Krichinskys, Kutuzovs, Meshcherskys, Michurins, Shirinskys,
Scriabins, Suvorovs, Turgenevs, Yushinsky (from tribe Usun/Wusun/烏孫), Yusupovs, and many more. Now,
unbeknown to their Y-DNA marker, these names are held as Russian, and their origin by default as
Slavic, but the transposition of the ancient DNA line on the modern cultural identity and the
lingua franca of the state territories is misleading.
It appears that tracing the custom of the Right of first night in Europe invariably ascends
to the horse nomadic tribes of the Sarmatian circle: Vandals/Burgund/Bulgars and their kindred
tribes, peculiarly affecting the feudal customs in France, Germany, and England, but the roots of
the tradition penetrate much deeper, it was first recorded by Herodotus in Anatolia. In the slavery
societies of ancient agricultural states like Rome, Greece, Persia, and all the way to China, the
use of slaves for sex was a daily affair, not worth specific annalistic notation, but unwittingly
universally recorded in references to a "child of a slave or a concubine". |
| Modern reconstruction of Y-DNA map in Ptolemy's time |
Reconstruction of ethnic tribes according to Ptolemy |
 |
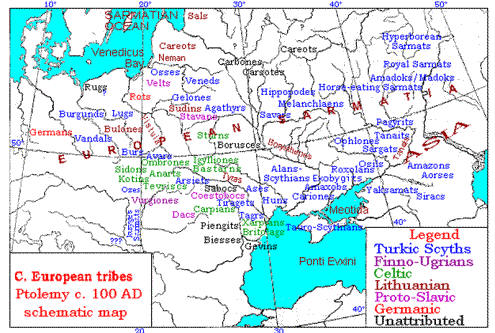
Virgiones of the map must be the same Burgundians ~ Bulgars per
Agathias;
Coestobocs of the map are held as Gauls/Celts;
Dacians are rated as Gepids, of the Goth/Vandal circle.
That leaves only Stavans in the upper Pripyat marches as tentative Slavs-Aryans. |
There is merit in comparing the Indian haplotypes with the R1b1a2 haplotypes — a group who
populates ~60% of Europe, living primarily in the British Isles, Spain, France, Belgium, Germany,
the Netherlands, and other Central and Western European countries. The typical ancestral haplotype
in R1b1a2 haplogroup, dated about 4,800 years before present, is as follows (Klyosov, 2011a):
13 24 14 11 11 14 12 12 12 13 13 29 — 17 9 10 11 11 25
15 19 29 15 15 17 17 — 11 11 19 23 15 15 18 17 36 38 12 12 — 11 9 15 16 8 10 10 8 10 10 12 23 23 16
10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12
There are 48 and 44 mutations between the above and the Indian R1a1 haplotypes shown earlier.
This formally places their common ancestor at more than 10,000 years before present and, in fact,
much earlier, at least 15,000 years ago. R1b1a2 bearers were not among the Aryans coming to India,
and it is very likely that they were (linguistically) not
Indo-Europeans then. Specifically, there is no supporting evidence that 4000 years before present
(ybp) bearers of R1b1a2 spoke Indo-European (IE) languages. On the other hand, Central Europe was
likely populated by R1b1a2 speakers of non-IE languages. Moreover, there are very few bearers of R1b
haplogroup in India, mostly on its Arabian Sea coast, and there were none of the R1b haplogroup
among the 367 tested Indian Brahmins (Sharma et al., 2009). Therefore, it is highly unlikely that
bearers of the R1b1 (as well as R1b1a2) haplogroup were among the Aryans, and, hence, they were not
among those carrying the Indo-European languages elsewhere in those times.
We are left holding two questions: first, from where did the R1a haplogroup arise and, second,
what was their migratory route that brought them to the
1) East European Plain (currently up to 62% R1a1, see above),
2) India and Iran (10% - 16% R1a1),
3) Anatolia (15% R1a1),
4) the Middle East (up to 7% - 13% R1a1), and
5) the Arabian Peninsula, where nowadays 2% - 10% of the population carries the R1a1 haplogroup
(Abu-Amero, 2009; Underhill et al., 2009)?
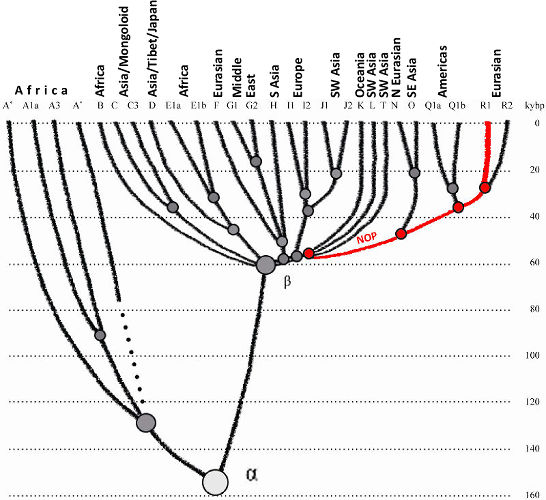
As described in (Klyosov & Rozhanskii, 2011), Europeoids (Caucasoids) appeared ~58,000 ybp. They
gradually branched to downstream haplogroups and migrated to the west, south and east. Haplogroup
NOP, which was among them, arose ~48,000 ybp, and moved eastward, presumably towards South Siberia
and/or adjacent regions. Haplogroup P arose ~38,000 ybp, apparently in South Siberia, and gave rise
to haplogroup R and then R1 ~30,000 - 26,000 ybp (see the diagram in Klyosov & Rozhanskii, 2011).
The timing of haplogroup R1a’s appearance can be reconstructed from series of R1a haplotypes, made
available from the databases (see the list in Materials and Methods and the Appendix). The most
ancient common ancestors of this haplogroup lived in: northern and northwestern China (in
particular, Xinjiang region (East Turkestan), which is the south
Altai area), in southern Siberia, in the Eastern Himalayas, India and Pakistan, the Comoros Islands,
and in Europe, where their bearers apparently migrated from the east during both the remote past and
later, for example, with the Scythians.
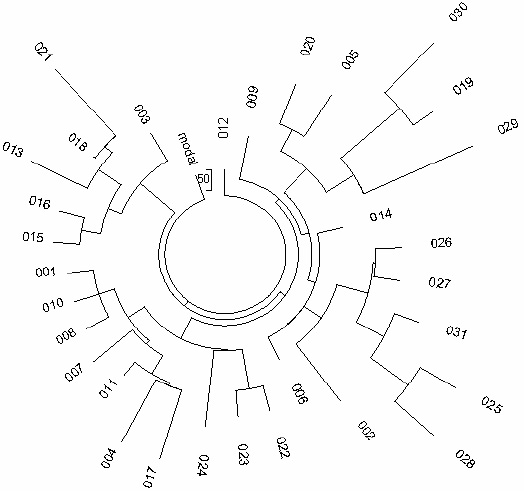
Northern China R1a Haplotypes
Apparently, the most ancient source of R1a1 haplotypes is provided by the people now living in
northern China (the area which historically was the ancestral territory of Uigurs, a
female dynastic line of the Huns. The Chinese colonial encroachment started during Qin dynasty, 3rd
c. BC). It was shown (Bittles et al., 2007) that for a number of Chinese populations,
such as Hui, Bonan, Dongxiang, Salars (all assembled primarily of Türkic
Uigurs), a percentage of R1a1 haplotypes reached 18% - 32%. Their haplotypes were not
provided in the paper, but the author, Professor Alan H. Bittles, kindly sent us a list of 31 of
five-marker haplotypes typed as R1a1, the tree of which is shown in
Figure 1. The haplotypes vary tremendously in their alleles, which already indicates that
their common ancestor lived in ancient times. For example, values of DYS19 varied between 14 and 17,
DYS388 between 12 and 14, and DYS393 between 10 and 13. It should be noted that mutations in the
last two markers occurred on average once in 4,545 and 1,320 generations, respectively. With a
correction for back mutations (Klyosov, 2009a) it occurs once in 8500 and 2400 generations. The 31
haplotypes contain 99 mutations from the deduced 5 marker base haplotype as shown here in the 12
marker FTDNA format with missing alleles indicated:
13 X 14 X X X X 12 X 13 X 30
This extent of mutations, which can be presented as 99/31/5 = .639 ± .064 mutation/marker, is a
very high value. Actually, it is a measure of how ancient a common ancestor might be (for a
comparison, contemporary European R1a1 and R1b1a2 haplotypes are separated by .250 - .270 mutations
from their common ancestors, see Klyosov 2011a, 2011b). It can also be presented as 99/31/.00677 =
472 →683 conditional generations; that is, it is 17,100 ± 2,400 years to the common ancestor of
these 31 haplotypes. The value of .00677 mutation/haplotype for conditional generation is the
mutation rate constant for the 5 marker haplotypes (Klyosov, 2009a).
2
Figure 1. The 5-marker haplotype tree for R1a1 haplotypes in Northern China.
The 31-haplotype tree was composed from data provided by Dr. A.H. Bitttles and collected in ethnic
communities Hui, Bonan, Dongxiang, and Salars (Bittles et al., 2007) (no haplotypes were provided in
the referenced article). A fraction of R1a1 haplogroup in said populations is 18%, 25%, 32%, and
22%, respectively (ibid). |
|
 |
Hui = Chinese designation for Moslems, from Hui =
Uigur; predominantly Uigurs with joined fractions of other Türkic and non-Türkic Middle Asian
extracts
Bonan (Bao'an=Uigur) = mix of Mongols, Hui, ubiquitous Han Chinese, and Tibetans, in Gansu;
Mongols, Hans, and Tibetans have nothing to do with the R1a, thus Bonan R1a ≈ Uigur;
Dongxiang (Bao'an=Uigur) = mix of Mongols, Han, Hui, and Tibetans, in Gansu, thus Dongxiang
R1a ≈ Uigur;
Salars = (Salar = Sary Uigur=Uigur) Oguzes + Uigurs, in Gansu.
With some nuances, these are people of Türkic genetic trunk.
On Tu, Xibe, and Tatars see notations below. |
Since these haplotypes descend from such an ancient common ancestor and contain numerous
mutations, this makes their deduced base (ancestral) haplotype rather uncertain. Therefore, the
quadratic permutation method was employed for the same set of haplotypes (Klyosov, 2009a). This
method does not require either a base haplotype or a correction for back mutations. The obtained
timespan is 19,625 ± 2,800 years to a common ancestor. This result is within the margin of error
with that calculated by the above linear method.
Therefore, haplogroup R1a arose at approximately 20,000 ybp with the territory geographically
belonging to Central Asia.
R1a1 Haplotypes from Altai
Thirteen Altai R1a1 haplotypes were listed in (Underhill et al., 2009), 12 of which showed a
rather recent base haplotype (the last marker is DYS461):
13 26 16 11 X X X 12 11 14 11 31 — 10
These 12 haplotypes have only 7 mutations per 120 markers from the above base haplotype, which
gives 7/120/.0018 = 32 →33 generations; that is, 825±320 years to a common ancestor. The same set of
the Altaian haplotypes in a different format is given in (Järve et al., 2009), with the base
haplotype (not listed in the cited paper)
13 26 16 11 11 17 X X 11 14 11 31
and the same 7 mutations per 120 markers. Therefore, this exactly correlates to the same timespan to
that of the common ancestor as given above. The same set of the Altaian haplotypes was given in
(Järve et al., 2009) in a significantly more extended format, with the base haplotype (the second
panel represents DYS 458, 437, 448, GATAH4, 456, 438, 594, 411S1 [two alleles], 596, 643, 645, 635,
YPenta1, YPenta2):
13 26 16 11 11 17 X X 11 14 11 31 — 15 14 19 11 15 11 8 10 11 9 10 8 23 11 10
All 12 haplotypes in the extended format collectively contain the same 7 mutations. In other
words, the added 15 "slow" markers did not produce mutations, and is an indicator of a quite recent
common ancestor of the haplotype dataset. However, this base haplotype differs by 6 mutations from
the base haplotype of the East European Plain, which in the same format is
13 25 16 11 11 14 12 12 10 13 11 30 — 10
and by 12 mutations from the East European Plain base haplotype in the extended format:
13 25 16 11 11 14 12 12 10 13 11 30 — 15 14 20 12 16 11 10 10 11 10 10 8 23 11 10
(a more extended 67 marker base haplotype of the East European Plain is shown below). These 6 and 12
mutations exactly fit the difference between the respective mutation rate constants for the two
haplotype formats, equal to .020 and .0404 mutations per haplotype per generation, respectively.
These mutation differences place a common ancestor of the Altaian and the East European Plain
haplotypes at 8100 ybp.
An additional Altaian haplotype from the list
14 24 17 11 11 15 X 12 12 10 13 11 31 — 15 14 20 13 15 11 10 10 12 10 10 9 24 11 8
differs by as much as 20 mutations from the above base Altaian haplotype, and by 12 mutations with
the East European Plain base haplotype. This places their common ancestors at 10,400 and 7300 ybp,
respectively.
R1a1 Haplotypes in Tuva
Four R1a1 haplotypes from Tuva, the region which borders with Altai in Southern Siberia, north of
Mongolia, were listed in (Underhill et al., 2009; Järve et al., 2009). Three of them are identical
in all their 26 markers
13 26 15 9 11 14 X 12 10 13 11 30 — 18 14 19 12 16 11 10 10 11 10 10 8 23 11 8
and significantly differ, by as many as 20 mutations, from the fourth one, which belongs to the
lineage found among the Altayans:
13 25 16 11 11 17 X 12 11 14 11 31 — 15 14 19 11 15 11 8 10 9 10 10 8 23 11 10
They also differ by 12 mutations from the East European Plain base haplotype. This places a
common ancestor of the Tuva R1a1 haplotypes at 10,000 ybp and also with the East European Plain base
haplotype by 7300 ybp.
What emerges from the analysis of the data is that the Altaian and the Tuva haplotypes have
apparently the same ancient R1a1 common ancestor, who lived 10,000 - 10,400 ybp. However, the
surviving DNA lineages, which "surfaced" only recently, particularly in Tuva, are different in Tuva
and in Altai, though all coalescent to said ancient common ancestor.
| We know of well-documented historical events that illuminate the demographical changes leading
to and in the early 1st mill. AD. These are consecutive relocations of the Türkic nomadic states of
Oguzes (740), Kangars/Bechens (colloquial Pechenegs, 750), Karluks (colloquial Karakhanids, 920),
Kimaks/Kipchaks (colloquial Polovetses, 990) from the greater Altai area to the west. Assuming that
2/3 of the population have relocated, 1/3 of the population minus military and hardship losses
stayed put, and carried on their genotype. The new demographic status at the destinations was a
blend of the previous population and the newcomers from the immediate east, linguistically also
Türkic, but with relatively higher proportion of the eastern Eurasian Y-DNA and mtDNA genes.
By the early 13th c., of the mid-1st mill. AD population, in the Altai probably remained only 10%
(1/32) of the early Altaian population, 90% of the early Altaians having moved to the
Middle Asia and Eastern Europe. At least partially, the outflow was replenished by the kindred
genotypes migrating from the east. In 1650 a Jungar-executed genocide reduced the number of Tele
people east of Tuva by ~90%, minority of whom found refuge in the Northern Altai (Altai in this
paper) and a majority settled in the Southern Altai (Tuva in this paper). |
R1a Haplotypes in the Eastern Himalayas
Five R1a1 haplotypes were listed in (Kang et
al., 2011), which showed a rather recent base haplotype (the last two markers are DYS437 and
DYS438):
3
13 25 15 10 11 14 13 14 10 13 11 30 — 14 11 11
These 5 haplotypes have only 4 mutations from the above base haplotype, which gives 4/5/.0215 =
37 →38 generations; that is, 950 ± 480 years to a common ancestor. However, the above base haplotype
has very unusual (for R1a haplogroup) alleles DYS426 = 13, and DYS388 = 14, and differs by 5
mutations with the East European Plain base haplotypes. This places their common ancestor at 6650
ybp. This is clearly a separate branch of ancient R1a haplotypes in Eastern Himalayas.
R1a Haplotypes in India and Pakistan
There are two principal sources of haplotypes
of haplogroup R1a in the Hindustan. One was brought by the Aryans in the middle of the 2nd
millennium BC, as it was described above, and supported below with more extended series of Indian
haplotypes. A timespan to the most recent common ancestor of these haplotypes varies between 4000
and 4600 ybp, and often around 4050 ybp, depending on a particular haplotype datasets. The base
(ancestral) haplotype of the Aryan (linguistically
Indo-European) haplotype in its 12 marker format is
13 25 16 10 11 14 12 12 10 13 11 30
This haplotype is nearly identical to that of the East European Plain base (see below), except
the latter came from a common ancestor who lived between 4,600 and 5,000 ybp as determined using
different haplotype datasets (Klyosov, 2009a; Klyosov, 2011b).
A more ancient source is presumably the South Siberian and/or Central Asian haplotypes brought to
the Hindustan during the westward migrations of R1a bearers between 20,000 and 10,000 ybp. Some
studies alleged that the most ancient common ancestors of R1a haplotypes were Indian; however, the
results were flawed by erroneous calculations of timespans using incorrect "population mutation
rates", which routinely converted the actual 3600 - 4000 ybp ("Indo-European" R1a1 in India) into
12,000 - 15,000 ybp. This was erroneously claimed as the proof of "origin of R1a in India."
Furthermore, high percentages of R1a in some regions in India or in some ethnic and/or religious
groups (such as Brahmins) were incorrectly claimed as the proof of the origin of R1a in India
(Kivisild et al., 2003; Sengupta et al., 2006; Sahoo et al., 2006; Sharma et al., 2009; Thanseem et
al., 2006; Fornarino et al., 2009). The application of the flawed approach resulted in confusion
amongst researchers in the field of human population genetics over the last decade. The course of
research is hopefully corrected by the application of today’s most recent developments of DNA
genealogy, which utilizes a principally different methodology (Klyosov, 2009a, 2009b, 2009c;
Rozhanskii and Klyosov, 2011; Klyosov, 2011b).
Forty-six of 6 marker R1a1 haplotypes of three different tribal population of Andra Pradesh,
South India (tribes Naikpod, Andh, and Pardhan) listed in (Thanseem et al., 2006) and shown in the
haplotype tree in Figure 2, contain 126 mutations; that is .457 ± .041 mutations per marker
(the mutation rate constant equals .0123 mutation/haplotype/generation, Klyosov, 2009a). It gives
7200 ± 960 years to a common ancestor (TMRCA = “Time to Most Recent Common
Ancestor” ). The base (ancestral) haplotype of those south Indian populations in the
FTDNA format is as follows:
13 25 17 9 X X X X X 14 X 32
This differs from the north-Indian "Indo-European" haplotype (see above) by four mutations on six
markers, which places their most recent common ancestor to approximately 11,600 ybp.
The ancient north China R1a1 base haplotype (see above) differs from the Andra Pradesh R1a1 base
haplotype by at least 5 mutations on the 5 available markers, which places their common ancestor at
approximately 22,000 ybp. Within a margin of error, it can be deduced that this is the same common
ancestor of the north China haplotypes. This mutational difference neatly fits the chronology and
direction of the migration, which continues from the ancient (non
linguistically
Indo-European) Indian haplotypes to the (linguistically)
Indo-European Indian haplotypes with their common ancestor (non-IE and IE) who lived approximately
11,600 ybp. Also, this data dovetails with the timing of the follow-up migration of R1a1 bearers
from Hindustan via Asia Minor (with a detection of the proto Indo-European language in Anatolia with
estimated divergence time of 9400 - 9600 - 10,100 ybp, see Gray & Atkinson, 2003; Renfrew, 2000;
Gamkrelidze & Ivanov, 1995) to Europe (with the arrival 10,000 - 8000 ybp, see below) and then to
the East European Plain (5000 - 4800 ybp, see below).
The analysis of this data and of these findings essentially unites most, if not all, concepts of
the "origin of Indo-European language" which have, at various times, placed the "origin" from India
to Iran, Anatolia, the Balkans, to the Russian Steppes (Gimbutas, 1973, 1994; Mallory, 1989; Dixon,
1997; Anthony, 2007), except that they were related not to the "origin," but to the passing areas of
the R1a1 migration.
Population geneticists typically mix DNA lineages and branches in their analysis whereby "phantom
common ancestors" emerge. This is exemplified with 110 of 10-marker R1a1 haplotypes of various
Indian populations, both tribal and Dravidian and Indo- European castes, listed in (Sengupta et al.,
2006). The resulting mixed haplotype tree is shown in Figure 3. It contains 344 mutations,
which is .313 mutations per marker, and results in a "phantom" 5275 years to a "common ancestor,"
just between the shown above 7180 ± 960 years for non-IE and 4050 ± 500 IE Indian haplotypes.
Figure 2. The 6-marker haplotype tree for R1a1 haplotypes in Andra Pradesh (tribes Naikpod, Andh,
and Pardhan), South India
(TMRCA 7200 ± 960 years).
The 46-haplotype tree was composed from data listed in (Thanseem et al., 2006). The designations of
haplotypes are those used in the article. |
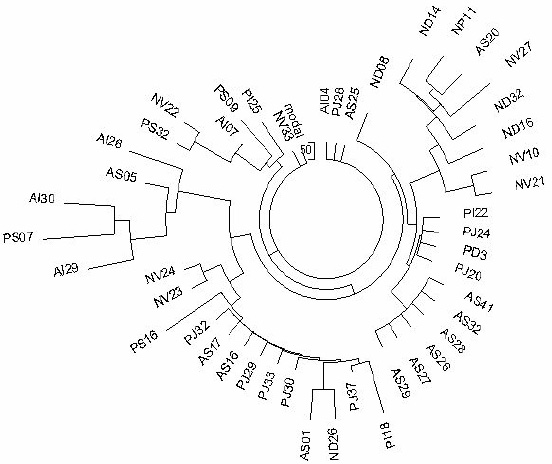 |
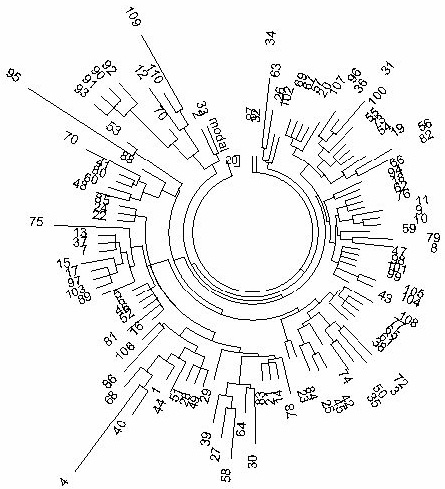
Figure 3. The 10 marker haplotype tree for R1a1 haplotypes in India (mixed population, including
tribes and castes) (TMRCA ~4050 years).
The 110-haplotype tree was composed from data listed in (Sengupta et al., 2006). The article
contains 114 Indian R1a1 haplotypes, however, four of them were incomplete. |
 |
4
For a comparison, consider the Pakistani R1a1 haplotypes listed in the Sengupta (2006) paper (Figure
4). Forty-two haplotypes contain 166 mutations, which gives .395 ± .031 mutations per marker,
and 6800 ± 860 years to a common ancestor. This value fits within margin of error to the
"south-Indian" 7200 ± 960 ybp; however, the base (ancestral) haplotypes differ significantly. The
base Pakistani haplotype is as follows (in the FTDNA format plus DYS461):
13 25 17 11 X X X 12 10 13 11 30 — 9
It differs from the south Indian "non-IE" and the north Indian "IE" base haplotypes by four and
two mutations on six markers, respectively. This places a common ancestor of the Pakistani and the
south Indian "non-IE" R1a1 populations at approximately 12,980 ybp which is within margins of error
with the 11,600 ybp reported above as the migration time through the Hindustan westward. The two
mutations place a common ancestor of the Pakistani and the "Indo-European" Indian populations more
recently, at 7800 ybp. This chronological trend might also point in the direction of the ancient
migration of R1a1 westward.
A more detailed consideration of the Pakistani R1a1 haplotypes, including separate calculations
of each of the four branches in Figure 4, results in a timespan of 8650 years to a common
ancestor for all of these branches (Klyosov, 2010a). In all, it does not change the principal
conclusions of this section.
R1a1 Haplotypes in Central Asia
Ten 10-marker Central Asian haplotypes were listed
in (Sen-gupta et al., 2006). They contain 27 mutations from the base haplotype
13 25 16 11 X X X 12 10 13 11 31— 9
which gives .270 ± .052 mutations per marker, and 4300 ± 940 years to a common ancestor. It is the
same value that we have found for the East European Plain and "Indo-European" Indian base R1a1
haplotypes.
Both the Central Asian base haplotype and the dating of a common ancestor described above are
supported by the latest data on extended 67 marker haplotypes that were collected in November 2011
in the R1a1 FTDNA Project. The Central Asian base haplotype was as follows:
13 25 16 11 11 14 12 12 10 13 11 31 — 15 9 10 11 11 24
14 20 32 12 15 15 16 — 11 11 19 23 15 15 18 19 34 38 14 11 — 11 8 17 17 8 12 10 8 11 10 12 22 22 15
10 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13
This is identical to the above-mentioned Central Asian 10- marker base haplotype in all the given
alleles. A common ancestor for the series of the extended haplotypes lived 3650 ± 590 ybp. The
Central Asian base haplotype differs from the East European Plain base haplotype (Rozhanskii &
Klyosov, 2009) by only 4 mutations in the 67 markers. This separates them in terms of the time of
their common ancestors
13 25 16 11 11 14 12 12 10 13 11 30 -- 15 9 10 11 11 24
14 20 32 12 15 15 16 — 11 11 19 23 16 16 18 19 34 38 13 11 — 11 8 17 17 8 12 10 8 11 10 12 22 22 15
10 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13
by only 4/.12 = 33 →34 conditional generations; that is by ~850 years.
The result is compelling and provides an exact fit with the expected migration pattern of the
R1a1 haplogroup from the East European Plain (~4600 - 4400 ybp) to Central Asia (3650 ± 590 ybp) on
their way to the South Urals and to the Hindustan.
What these findings suggest is that there are two different subsets of the Indian R1a1
haplotypes. One was brought by European bearers known as the Aryans, seemingly on their way from the
East European Plain through Central Asia in the middle of the 2nd millennium BC. The other was much
more ancient and migrated from South Siberia/northern China to India 12,000 years ago. This
migratory wave continued through the Iranian Plateau westward (via Anatolia and the rest of Asia
Minor), to the Balkans and then further into the European continent.
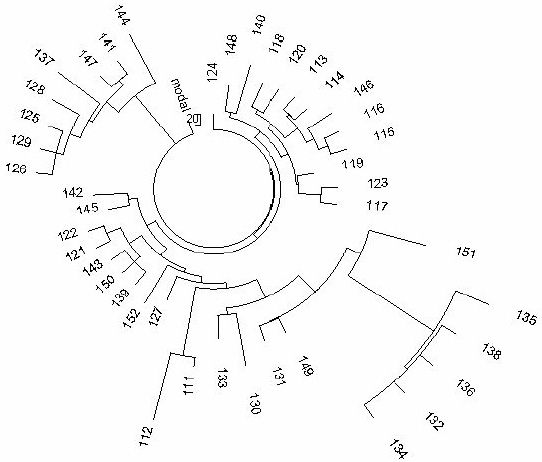
Figure 4. The 10-marker haplotype tree for R1a1 haplotypes in Pakistan.
The 42- haplotype tree was composed from data listed in (Sengupta et al., 2006). |
 |
5
R1a1 Haplotypes of the Comoros Islands
Fifteen R1a haplotypes have been found among 381 tested men on the Islands. Three of them were
R1a*-SRY10831a, and twelve were R1a1 (Msaidie et al., 2011). The cited study did not generate any
chronological estimates based on the haplotypes, and considered only 8 marker haplotypes (for a
typical "population genetics" analysis without separation of haplogroups) while, in fact, determined
17 marker haplotypes.
The base haplotype for said 12 R1a1 haplotypes is as follows:
13 24/25 15 11 12 14 X X 10 13 11 18 — 16/17 14 19 12 15 11 23
All have 104 mutations; that is, .51 ± .05 mutation per marker. This high value points at a
significantly more ancient common ancestor compared with that in the East European Plain, Central
Asia and the Indo-European Indian R1a1 populations (.28, .27, .24 mutations per marker,
respectively). Furthermore, it is more ancient compared with the old south Indian and Pakistani R1a1
populations (.457 and .395 mutations per marker, respectively, see above). Indeed, a common ancestor
of the Comoros R1a1 haplotypes lived .51/.02 = 255 →340 conditional generations; that is 8500 ± 1190
ybp.
For a comparison, the East European Plain base haplotype in the same format is as follows:
13 25 16 11 11 14 X X 10 13 11 17 — 15 14 20 12 16 11 23
It differs by as many as 7 mutations from the Comoros base haplotype. This places their common
ancestor at 9900 ybp. It is reasonable to suggest that this common ancestor was one of those R1a1
who were moving westward along the Iranian plateau and Asia Minor almost 10,000 ybp. Indeed, the
dating around 9900 ybp is rather typical for archaeological settlements in Asia Minor with known
dates of 10,200, 9900 and 9000 ybp (Myres et al., 2010). It is not necessarily true that the bearers
of R1a1 were in the Comoros Islands 9900 ybp since it is known that the Persian traders had expanded
their maritime routes to Madagascar by 700-900 AD (Msaidie, 2011).
|
The reference to archaeological settlements in Asia Minor at around 10,000 ybp is interesting,
not because the cited dates appear not to appear in the Myres et al., 2010
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3039512/, but because the passing through the Asia
Minor R1a1 hunter-gatherers were unlikely to leave archaeological traces other than their campsites
and pre-Kurgan burials. The hunter-gatherer economy did not encourage permanent locations with
permanent structures like those among entrenched agriculturists, but they tended to follow seasonal
cycles harvesting known products in known ranges. Fertile forests could reduce the scale of
transhumance, but the absence of wheel transport, draft or riding animals still necessitated
traditional technology of quick and light temporary housing along the lines of the native American
wigwams, practically undetectable archaeologically. The dating around 9900 ybp is typical for
permanent agricultural archaeological settlements in Asia Minor, not for the roaming
pre-agricultural and pre-animal husbandry hunter-gatherers. And since no soul on this planet could
write before the invention of literacy, any notion of the of R1a1 hunter-gatherer language around
10,000 ybp is rather speculative. It appears that allowing liberally a timespan from 10,000 to 1100 ybp for the arrival of R1a1 on
the Comoros conceptually conflicts with the rigid postulated arrival of the R1a1 in the Eastern
Europe. |
R1a1 Haplotypes in the Arabian Peninsula
Sixteen R1a1 10 marker haplotypes from
Qatar and United Arab Emirates were published (Cadenas et al., 2008). They split into two branches,
and their base haplotypes
13 25 15 11 11 14 X Y 10 13 11 30
13 25 16 11 11 14 X Y 10 13 11 31
differed by only one mutation. Their common ancestor lived 3750 ± 825 years bp. Since a common
ancestor of R1a1 haplotypes in Armenia and Anatolia lived 4500 ± 1040 and 3700 ± 550 years bp,
respectively (Klyosov, 2008), the three dates do not conflict with each other. They were not part of
the ancient migrations of 12 - 9 thousand ybp, but they were most likely on the military expeditions
of the (Aryan) R1a1 from the East European Plain southward through Anatolia, Mitanni, and to the
Middle East and the Arabian Peninsula around 4000 - 3600 ybp. As mentioned earlier, today there are
between 3% and 9% of R1a1 in those regions, among them members of famous tribes such as
Quraish/Quraysh (Muhammad, the founder of the religion of Islam, was born into the Quraysh tribe),
Al Tamimi (Banu Tamim) and others.
|
We know nothing about military expeditions of the Aryans from the East European Plain southward
through Anatolia, Mitanni, and to the Middle East and the Arabian Peninsula around 4000 - 3600 ybp.
What is known is that Arians did not ride mounted into India, they did not bring horses to India,
they rode oxen carts. Which means that the Aryan military expedition from the East European Plain
southward to the Arabian Peninsula around 4000 - 3600 ybp went along the lines of the much later
Darius 516 BC march against Scythians, with endless columns of foot soldiers, and oxen-pulled wagons
and supply trains. The idea of such a lethargic Aryan expedition to the far-away Arabia after the
Arabs mastered the use of the camels appears to be quite a figmentary construct. Bands of randomly
roaming not numerous clans in search of better farmland, like an Aryan version of the slightly later
father Abraham, seems to be more appropriate for the 4000 - 3600 ybp times. The alternate candidates would be the East European Tiber Grave Kurganians, who would not only
bring along their R1a1 allele, but also horses, kurgan burials with horses and travel supplies,
hand-made ceramics tempered with shells, and expressly non-mediterranean anthropological remains,
technologically and culturally predating Scythians, but carrying unmistaken traces of their kinship.
Unfortunately or fortunately, the earliest monuments of the Scythian-circle incursions in the Near
East are the traces attributed to the Cimmerians, dated by about 3000 ybp; but according to
A.Ivanchik(2001), the Cimmerians and Scythians are archeologically indistinguishable, and we have no
paleogenetic information on them. According to A.Klyosov, the Tiber Grave Kurganians should leave
behind their predominant R1b alleles, not the R1a1. |
Much more reliable data is obtained with extended 67 marker haplotypes from the Arabic FTDNA
project. Twenty-seven haplotypes from Qatar, Kuwait, Saudi Arabia, UAE, Oman, Bahrain and Syria form
a separate branch on the haplotype tree and result in the following base haplotype:
13 25 16 11 11 14 12 12 10 13 11 30 — 15 9 10 11 11 24
14 20 32 12 15 15 16 — 11 11 19 23 15 16 18 19 35 38 13 11 — 11 8 17 17 8 12 10 8 11 10 12 22 22 15
10 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13
Therein 499 mutations exist and 499/27/.12 = 154 →182 generations; that is, 4550 ± 500 years from
a common ancestor of the Arabic haplotypes — practically the same as that for the East European
Plain R1a1 common ancestor (see above). The two differ by only 1.4 mutations in all the 67 markers;
that is, 1.4/.12 = 12 generations apart, a ~300 year difference between the Russian Plane R1a1
common ancestor and the Arabic haplotypes common ancestor. The exception being that the Arabic
haplotypes are typically coupled with the downstream L342 SNP mutation. The difference places their
common ancestor at ~4825 ybp, which is the East European Plain base (ancestral) haplotype. This is
the same Aryan haplotype that was brought ~4500 ybp from the East European Plain in a star-like
manner to India, Iran, Anatolia, the Arabian Peninsula to arrive there a thousand years later, in
the middle of the 2nd millennium BC.
|
The refugees from the Central Europe that fled to the East Europe from a pogrom inflicted by the
R1b Kurgan invaders might have developed their lingua franca right there, in the Eastern Europe,
aggregating numerous vernaculars of the refugee crowds and the preceding local tribes. The time
around 4500 ybp present was one of most swirly times in Europe, the R1b Kurganians went from the
Eastern Europe west to Europe, and the R1a farmers fled to the Eastern Europe. That is, if if if we
accept the conjecture that the appearance of the 10,000-years old R1a alleles in the Balkans was a
jump from the Anatolia 9000 ybp, and not a result of the well-documented Kangar/Bechenek mass
migration to the Balkans in the 700-1000 AD carrying the Middle Asian 10,000-years old R1a alleles
to the Balkans. If we accept the Anatolian R1a migration, and then accept the Balkan R1a migration,
we would get a Eastern European melting pot with the bulk of the old R1b terrible East European
Kurganians and an inflow of the scared and destitute Central European farmers. The farmers, though,
did not stay around for too long with the Kurganians, they fled once again in search for safer
grounds, carrying their newly formed syncretic culture and the emerging Indo-European language
southward in a star-like manner. The corollary of the suggested whirlwind scenario is that the newly formed language has to bear
marks of creolization, a loss of most of the grammatical attributes, and a development of
analytic-type language that primarily operates with the roots of the words, like English and
Chinese. The creolization would have taken place independently of the location, be it in an
intermediate refuge in the Central Europe, or in the later refuge in the Eastern Europe.
Another corollary of the suggested whirlwind scenario is the effect of the bottleneck, invariably
created during any migration. Only a part of the available gene pool is transplanted to the new
location. |
Recent developments in the phylogeny of R1a1 haplotypes coupled with the DNA genealogy analysis
have shown that the migrations of R1a1 from the East European Plain in the described star-like
manner were accompanied with the R1a1-L342 (around 4400 ybp) and then its downstream L657 subclade.
The L342 subclade is almost absent on the East European Plain, and it appears in the Bashkir
population in the east, in Kazakhstan (L342 →L657) south-east, in India (L342 →L657), and in the
Middle East (including the Arabian Peninsula, L342 →L657). It shows primary directions of the Aryan
(R1a1) migrations after ~4800 ybp.
The Arabian R1a1-L657 haplotypes along with all known Iranian, Indian and Kazakh L657 haplotypes
have the following L657 base haplotype:
13 25 16 10 11 14 12 12 10 13 11 31 — 16 10 10 11 11 24
14 20 32 12 15 15 16 — 11 11 19 23 15 16 18 19 35 40 14 11 — 11 8 17 17 8 12 10 8 11 10 12 22 22 15
10 12 12 14 8 13 23 21 12 12 11 13 11 11 12 13
Its common ancestor lived 3000 ± 400 ybp. The above base haplotype differs by 9.85 mutations from
the East European Plain base haplotype (some mutations are fractional ones), which places the
R1a1-L657 and R1a1 East European Plain common ancestor at 5000 ± 600 years bp.
|
The area of Bashkiria served as refuge area for very diverse ethnic groups and for as long as we
have records on Bashkirs. Nowadays, Bashkirs are a minority in their own country, < 30%, so anything
from anywhere can be found there. Besides Bashkirs, in Bashkiria and surrounding areas live Bechens,
Abdaly/Ephtalites, Kipchaks, Huns, probably the same
Seklers/Szeklers/Izgils/Ishkil/Ichgil/Äsägel/Askel/Askil//Esgil/Esgels/Esegels/Esegs called Esgels
in the Tatar-Bashkir area, “Hermihions/Hionites/Red Chions” in Persian, Ch. Asitsze/Pin. Asijie,
Sijie 思結/Hermihions, and probably other tribes. The Kazakhstan's south-east was a territory of
Usuns, with remnants of As-Tokhars and Saka, and probably with some Kangars. Nowadays, it is the
territory of the Senior Juz, which includes the oldest annalistically known Türkic tribes. That
territory was not as heavily colonized as the northwestern Kazakhstan, but it still suffered all
genetic consequences of colonization, especially in the last half-century, and again anything from
anywhere can be found there in meaningful proportions.
Seklers/Szeklers/Izgils/Esgels/Esegels/Eseg was a dynastic tribe of the Scythians and their late
(and leftover) state in Dobruja; they were one of the triumvirate leading tribes of the Bulgars,
known as Esegs, with autonomous status; in the 10th c. in Bulgaria was a coup later repeated by Ivan
the Terrible, the hereditary possessions were converted to provinces, governors appointed, and only
those who accepted demotion were appointed as governors of their provinces without a right to
succession; after that the eastern Esegs disintegrated and disappeared as a nation; the western
Esegs under a Hungarian name Szekler ([E]sek + lar [Tr. pl.]) continued autonomous existence as an
allied nation supplying troops for the suzerain; that's how the dynastic tribe of the Scythians
became Hungarian service cossacks. In the Chinese annals the tribe Ezgil/Ezgel (Ch. Asitsze) was
described as strongest tribe of the Eastern Hun confederation, it later became a maternal dynastic
tribe in the Türkic Kaganate. In Ezgils we see the Ases of Tacit and Strabon, they were a dynastic
tribe of the Tokhars, conquerors of Bactria, and Ash-guzai of the Assyrians. The "service class" is
a great understatement. If their haplotype was the "Atlantic modal haplotype", then Ases and Seklers
are known from Pacific to Atlantic, appearing everywhere as a dynastic tribe; Chingiz-khan was from
the Uigur House of Djalairs, which were descendents of the Türkic Kaganate's maternal dynastic tribe
of Ezgils. And timewise Ases now extend from 4th millennium BC to today, see Yu.Zuev's work
http://s155239215.onlinehome.us/turkic/29Huns/Zuev/ZuevStrongTribeEn.htm. It is very exciting to
locate not only the Scythian royals, but the Ases/Tochars, mysterious Picts in conical hats, and way
beyond to the "Atlantic modal haplotype", it is like a dream. Picts were building huge corrals out
of volcanic tuff, the corrals are still standing.
There is more. The Transylvania and Dobruja Seklers maintained their traditional links until the
catastrophe in ca 1920, when they were kicked out from their land into a foreign country. With their
traditions, and incest prohibitions, they had a permanent martial partner, which supplied wives for
their tribe. In the old days, from before our era into the Middle Age, their permanent martial
partner was the Tokhar tribe. The alliance could break from time to time, but because they stayed in
Transilvania and Dobrija for 2 millennia, there is a high chance that they had just one martial
partner for 2 millennia, and older people still know who that is, because incest laws were strictly
observed till the disintegration of the rural society, which happened only very recently. |
R1a1-L342 Bashkir and Szekely/Seklers (Hungarian) Haplotypes
As noted in the
preceding section, migrations of the ancient Aryans eastward (and in some cases westward, as
illustrated below with the Szekely L342 R1a1 haplotypes) have resulted in the appearance of the
downstream R1a1 subclades, such as L342, among the Bashkirs. The respective L342 base haplotype is
as follows:
6
13 24 16 11 11 15 12 12 12 13 11 31 — 15 9 10 11 11 24
14 20 31 12 15 15 15 — 11 10 19 23 16 15 19 19 35 38 14 11 — 11 8 17 17 8 12 10 8 11 10 10 22 22 15
10 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13
Their common ancestor lived only 1300 ± 250 ybp; however, the base haplotype differ by as many as
14 mutations from the East European Plain base haplotype. This places their common ancestor, for the
Bashkirs and the East European Plain, at 4700 ± 500 ybp. This is again the Aryan R1a1 common
ancestor on the East European Plain.
The anchor points are
1. Indian upper castes now have Y-DNA (72%), now also found predominant in the East Europe, with a
common ancestor at 4500 ybp;
2. In Bashkiria now was found a line of that common Y-DNA with common ancestor at 1300 ybp, a
hallmark of the past bottleneck;
— Consequently,
1. Some 1300 years ago, at 700 ± 250 AD, or in the period of 450 to 950, some refugee came to
Bashkiria and settled there;
2. Or later a group of migrants moved to Bashkiria after experiencing in that period a bottleneck
elsewhere;
3. During that period,
3.1 Hunnic Empire disintegrated (ca 450 AD), and dispersed east, possibly leaving a Y-DNA trail in
Bashkiria (e.g. Esegs/Seklers);
3.2 Ephtilite Empire disintegrated (ca 550 AD), and dispersed west and north-east (Imenkov Culture)
3.3 Great Bulgaria disintegrated (ca 660 AD), and dispersed west and north-east
3.4 Kangar Union south of the modern Bashkiria disintegrated (ca 550 AD), and dispersed north-west
(Bechens/Patsinaks/Pechenegs/Bosnyaks)
3.5 Oguz Yagbu state expanded to Eastern Europe (ca 760 AD), becoming an active local participant,
with later disintegration
3.6 Khazar Kaganate disintegrated (ca 950 AD), and dispersed to the west and north-east
3.7 Start of Kipchak/Kimak advance westward (ca 950 AD), displacing and absorbing local tribes
4. All these Türkic entities and their constituent tribes could have seeded the Bashkir refugees;
5. Numerous clans of Indian upper castes derive their origin from the Saka/Scythians, Hunas and
Abdaly/Ephtilites, who could seed both India and Eastern Europe with their Y-DNA without any
assistance of the Indo-Aryans.
6. The common ancestor at 4500 ybp could have arose anywhere in the Middle or Central Asia, and
spread their seeds in various areas of Eurasia. |
There is quite a distant L342 lineage among descendants of Hungarian Szekely servicemen, recorded
in the first 1602 military census. The lineage is only 675 ± 260 years "old". However, its base
haplotype
14 23 17 11 11 14 12 12 10 14 11 32 — 17 9 10 11 11 24 14 20 31 12 14 15 15 — 11 12 19 23 16 15
20 19 34 38 13 11
contains as many as 15 mutations in the first 37 markers from the respective L342 Bashkir base
haplotype. This places their common ancestor to 3500 ± 400 ybp. It apparently reflects migrations of
R1a1-L342 bearers from the Ural region westward to Transylvania along with Finno-Ugric migrations of
those times.
|
Knowing the dynastic conservatism of the Türkic tribes, it is more plausible that Esegs/Seklers
accompanied the Northern Huns to Dzungaria in 93 BC, and then from Dzungaria to Altai in 126 AD and
on to the Aral area that extended their pastures to the Kama river and Bashkiria. By 334 Bulgars
(Hunno-Bulgars) are known in the Azov area, by 337 Huns become known to the Classical writers. The
female dynastic tribe of the Huns, although not specifically known to the Classical writers, not
only accompanied Huns, but also left numerous traces of their estates called Hatun, Hotyn (Queen),
and the like in the Eastern Europe. It is known that after the Hunnic retreat from Pannonia, Huns
remain in their domains in Transylvania and Dobruja, which later were recorded as Sekler lands.
Today, Seklers constitute a largest minority in Europe, 3 mln people. With
all the political, religious and nationalistic redrawing of the map, Seklers had to leave noticeable
genetic traces in the modern Rumania. The notion of the Sekler migration from the Ural region
westward to Transylvania along with Finno-Ugric tribe(s) may be not far from the truth, allowing for
migration in few stages, first with the Huns in ca 350s (not documented), and then with Magyars to
Levedia (ca 750) and on to Atilkuzu (889) and on to Pannonnia (990) (documented). We still do not
know the affiliation of 3 Türkic Kubar tribes, who were led to Pannonia by Shilka's son Arbat (Hung.
Arpad) of the Dulo dynastic line, along with 3 Finno-Ugric Magyar tribes. |
R1a Haplotypes along the Ancient Migration Path from South Siberia to Europe
Three
principal studies have been published recently, that contain hundreds of R1a1 haplotypes from all
over the world (Underhill et al., 2009; Zhong et al., 2010; Shou et al., 2010). Analysis of those
haplotypes and the chronology of their common ancestors have not been undertaken by the authors of
these studies. Figures 5-7 show general views of R1a1 haplotype trees, that were calculated
from the data. The purpose for including pictures of these trees was not to analyze their fine
structure in detail (Klyosov, 2010a, 2010b), but to demonstrate their complex multi-branch
structure, hence, ancient origins. For example, relatively young trees (young "age" of their common
ancestor) are often rather symmetrical and relatively uniform, such as the East European Plain R1a1
haplotype tree with a common ancestor 4600 ybp (Figure 8).
Analysis of R1a1 haplotypes and their branches on the trees in Figures 5-7 shows that
their ancient common ancestors lived in south Siberia and Altai (belonging to both south Siberia and
Central Asia). Their ancient descendants carried the R1a1 haplogroup while migrating from North and
North-Western China, across Tibet and Hindustan, and then along the Iranian Plateau, from Asia Minor
and finally into Europe. Some remnants of ancient R1a1 were left in Cambodia, Nepal, Oman, Israel,
Iraq, Egypt, Crete, the Caucasus, Russia, Estonia (the respective haplotypes are recovered from data
published in Underhill et al., 2009, Zhong et al., 2010, Shou et al., 2010). Results of the dynamics
of mutation in these haplotypes significantly differ from those in the contemporary European R1a1,
except one ancient and distinct lineage of R1a1 in Europe (see below). Their common ancestors as
thusly reconstructed, lived from 20,000 ybp in south Siberia/northern China through 12,000 - 11,000
ybp in Hindustan and 6900 ybp in Uigurs in north-western China.
|
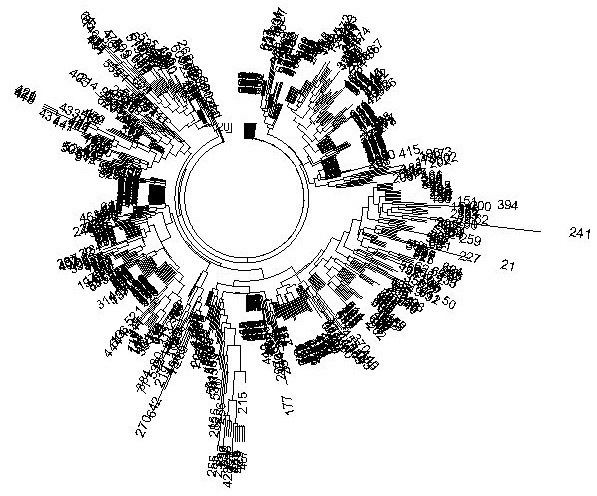
Figure 5. R1a1 10 marker 638-haplotype tree from all over the world, composed based on data
published by Underhill et al. (2009) |
 |
|
Figure 6. R1a1 8 marker 365-haplotype tree from all over the world, composed based on data
published by Zhong et al. (2010) |
 |
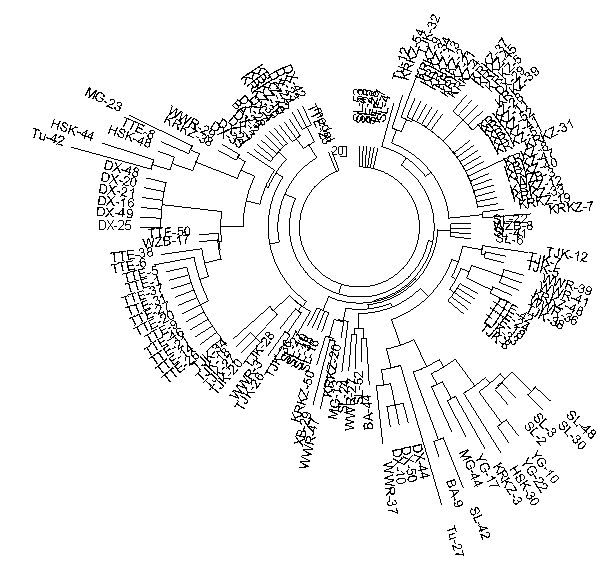
Figure 7. R1a1 8 marker 131-haplotype tree collected in North-Western China, composed based on
data published by Shou et al. (2010)
The lower right branch consists of R1* haplotypes |
 |
|
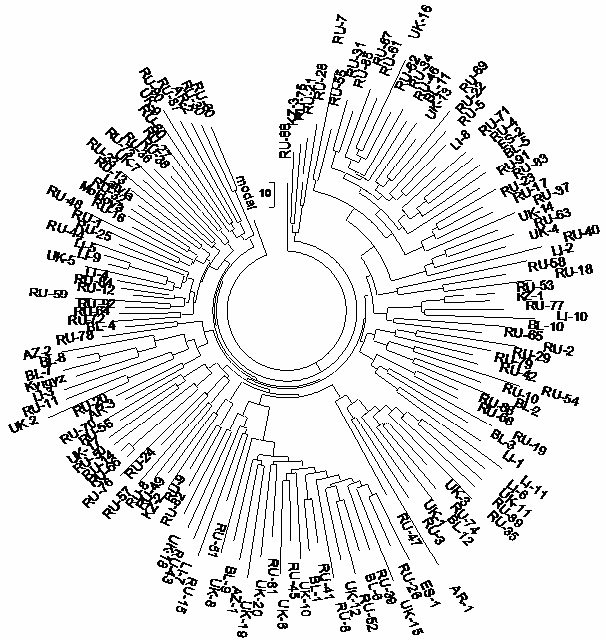
Figure 8. R1a1 67 marker 148-haplotype tree collected in Russia and Ukraine (East European Plain),
published by Klyosov (2011b) |
 |
8
Typically, ancient common ancestors are recognized by the distinct DYS392 = 13, unlike typical
DYS392 = 11 in most of European (and elsewhere) R1a1 haplotypes. The study by Underhill et al.
(2009) listed four Egyptian R1a1 haplotypes, two of them having DYS393 = 13, and two DYS393 = 11.
These four haplotypes have their common ancestor of ~13,275 ybp.
|
The great TMRCA ~13,275 ybp shows that they came from quite different lineages. In reverse
chronological order, Egypt, also called Dawla al-Turkiyya = State of Turkey, absorbed these
known waves of Türkic peoples: Ottomans (any of the 25 main Türkic tribes of Anatolia, 1500s-1800s),
Mamluks (mostly Kipchaks, 1200s-1400s), Saracenes (Bulgars, Savirs, Kipchaks, Kimaks, 700s-1100,
demographically minor). |
The very top of the tree (Figure 6) contains 18 base haplotypes, which are identical to
each other, and expressed in the 9 marker format as follows:
13 25 16 11 X X X 12 10 13 11 30
In this particular case these identical haplotypes are from Russia, Turkey, Ukraine, Slovakia,
Iran, Nepal, India, and Hungary (Compare the location of the Türkic states
in the last 2 millennia: Russia, Turkey, Ukraine, Balkans, Iran, India, Hungary, and China).
The short haplotype format does not allow them to be resolved any further, but with the available 9
markers this base haplotype is an exact (albeit partial) reproduction of the base haplotype of the
East European Plain. Furthermore, the tree in Figure 8
produces exactly the same 67 marker base haplotype of the East European Plain. The whole tree
contains 148 haplotypes with 2748 mutations from the base haplotype. It produces 2748/ 148/67 = .277
± .005 mutations per marker, and .277/.12 = 155 →183 generations; that is 4575 ± 470 years to a
common ancestor of the East European Plain base haplotype.
|
This general picture for what is called here the East European Plain base haplotype (R1a) parallels
the designation “Atlantic base haplotype” (R1b) for Western Europe, tentatively attributed to the
Türkic peoples. As it happens, among the Türkic peoples, R1a and R1b are predominating haplotypes. |
The whole pattern of ancient migrations of bearers of R1a1 haplotypes shows that after they had
arrived to Europe via Asia Minor, as it is described above, between 11,000 and 8000 ybp (see below),
they moved to the East European Plain in the beginning of the 3rd millennium BC. It coincided
time-wise with the arrival of bearers of R1b1a2 haplotypes in Europe. From there R1a1 split into
three principal streams. One stream migrated south, over the Caucasus to Anatolia, the Middle East
and the Arabian Peninsula. The second stream went eastward to South Ural, the Andronovo
(1800–1300 BC, 3800–3300 ybp) and Sintashta
(2100–1800 BC, 4100–3800 ybp)
archaeological cultures in the 2nd millennium BC, between 4000 and 3000 ybp, and then split into two
migration paths. One went south to India as the legendary Aryans, another went further east to Altai
and the Northern China. This closed the loop of the ancient migrations of R1a1. Yet the third stream
went south-east to the mountainous terrain of Middle Asia in ~4000 ybp, and some 500 years later
moved to Iran, otherwise known as the "Avesta Aryans."
|
Neither Andronovo nor Sintashta are viewed by third parties as incontrovertible Indo-Iranians or
ancestors of the "Avesta Aryans". The popular references to the "Russian scholars" or "Russian
school" are misleading, since the post-Soviet Russia has developed, in addition to the official
school, fairly independent schools that cautiously or openly laugh at the frills of the official
postulastes. If anything, the attribution should be rated controversial or disputable. Among the
main points disputing the Indo-Iranian linguistic attribution of the Andronovo and Sintashta are:
1. The ethnology of the nomadic Andronovo and Sintashta is not compatible with the "legendary
Aryans" Indo-Iranian agricultural ethnology.
2. NO archaeological evidence exists for any variant of the Andronovo culture either reaching or
even influencing the cultures of Iran or northern India in the second millennium. Not a single
artifact of identifiable Andronovo type has been recovered from the Iranian Plateau, northern India,
or Pakistan (Bryant E., Patton L., eds. "The
Indo-Aryan Controversy: Evidence and Inference in Indian History", Routledge, 2004, p 155).
3. The nomadic Andronovo and Sintashta burials are Kurgan burials with travel inventory accompanying
the deceased. The "legendary Aryans" did not bring Andronovo-type Kurgan burials to the Iranian
Plateau, India, or Pakistan.
4. NO zooarchacological evidence exists for the presence of the horse in Iran until the last
centuries of the second millennium BC and even then such finds are exceedingly rare. In South Asia
the first appearance of the horse is at Pirak. Pakistan, and dated to c. 1700 BC (Jarrige and
Santoni 1979) (Bryant E., Patton L., eds. "The
Indo-Aryan Controversy: Evidence and Inference in Indian History", Routledge, 2004, p 155).
5. By the time of the nomadic Andronovo and Sintashta, the mounted horse riding in Kazakhstan, in
the Andronovo area, was around for 2.5 thousand years, but the "legendary Aryans" did not bring
mounted horse riding to India and Iran.
6. Dating of the Andronovo culture, with respect to the chronology of its geographical distribution
and cultural variation is non-existent. Chronological sequence is “floating”, and so is the
careful explication of the cultural variants of the Andronovo. Researchers focus upon a single
attribute, without considering temporal or typological variations. Typological parallels are drawn
in the absence of chronological control. Chronological synchronisms are made on assumed
typological parallels (Bryant E., Patton L., eds. "The
Indo-Aryan Controversy: Evidence and Inference in Indian History", Routledge, 2004, p 156).
The circular logic rules.
7. The nomadic "legendary Aryans" did not bring lactase tolerance to either Iran, India, or China,
incredible for people living on a diet of milk and meat.
8. Three arguments are advanced on the identity of the Andronovo culture with the Indo-lranians:
(I) they are “in the right place at the right time." This argument remains thoroughly unconvincing;
(2) parallels between the material culture and environment of the Andronovo vs. commentaries in the
Rigvcda and Avesta are taken to confirm the Indo-Iranian identity of the Andronovo. The parallels
are far too general to offer confidence in these correlations;
(3) later Scythians (Saka), known to be Iranian, occupy the same territory and share generalized
similarities in material culture with the Andronovo. Thus, the ancestral Andronovo must be
Indo-Iranian (Bryant E., Patton L., eds. "The
Indo-Aryan Controversy: Evidence and Inference in Indian History", Routledge, 2004, p 156).
Since later Scythians (Saka) are nowhere close to Iranians, and are positively known to be Türkic in
the eyes of the contemporary Eastern and Western witnesses and outside of recent Eurocentric
scholarship, the last argument is self-defeating. Softly stating, all three arguments are thoroughly
unconvincing.
However, the credulous attitude of the authors is irrelevant to the authors' findings on the R1a
march in time and space.
|
R1a1 Haplotypes of the Current Descendants of the Legendary Aryans in India
The
R1a1 FTDNA Project in November 2011 contained 101 Indian haplotypes. Their base haplotype in the 25
marker format
13 25 16 10 11 14 12 12 10 13 11 30 — 16 9 10 11 11 24 14 20 32 12 15 15 16
contained only 1.4 mutations on average, compared with the East European Plain base haplotype (see
above). This translates into 1.4/.046 = 30 →31 generations, or ~775 years between their common
ancestors. In terms of time, this is a close distance between the East European Plain and the Indian
base haplotypes, and it fits with the time spans for the East European Plain R1a1 common ancestor
(4600 - 4800 ybp) and the Indian common ancestor (~4050 ybp), determined independently. This is the
historical Aryan base haplotype.
|
Probably, for a country of 1,2 billion people, 101 samples is not a representative measurement, if
it was for the upper castes only it would not ne a representative sample, and even if it was for the
Brahman caste only it would give only a rough idea. The 101 sample with TMRCA ~775 years depicts a
bottleneck that occurred around 1200 AD, possibly outside of the modern Indian territory, and may
reflect, for example, an exodus of Horezmians after the Chingizkhan's takeover of Horezm and
ravaging its cities and fields, exodus resulting from the Chingizkhan's invasion of Punjab, Ulug
Khan army invasion, and the like. All these armies were drafted from Central and Middle Asia
subordinated to Chingizids. |
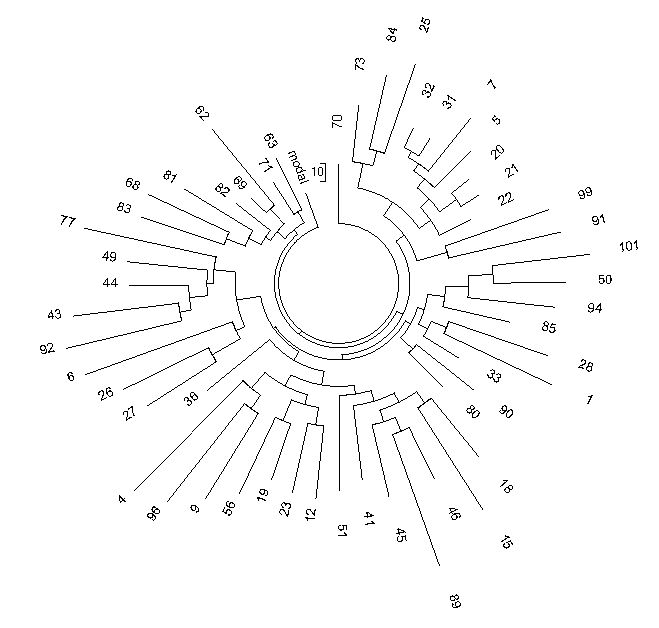
Figure 9 shows that the Indian R1a1 haplotype tree contains five principal branches. Their
base haplotypes in the 25 marker format are as follows (clockwise from the top):
13 24 16 10 11 14 12 12 10 13 11 30 — 15 9 10 11 11 23
14 20 32 12 15 15 16
13 25 16 11 11 14 12 12 10 13 11 30 — 16 9 10 11 11 24 14 20 32 12 15 15 16
13 25 15 10 11 14 12 12 10 14 11 30 — 16 9 10 11 11 25 14 20 32 12 15 15 16
13 25 16 10 12 14 12 12 10 13 11 31 — 16 9 10 11 11 24 14 20 32 12 15 15 16
13 25 16 10 11 14 12 12 10 13 11 31 — 15 9 10 11 11 24 14 20 32 12 15 15 16
Figure 9. R1a1 53-haplotype 25-marker tree collected in the Indian FTDNA Project database
(November 2011).
The Project contained 101 of 12 marker haplotypes, but only 53 of them were in the 25 marker
format. |
 |
8
A superposition of these five base haplotypes gives the above Indian "Indo-European", the Aryan
R1a1 base haplotype. All five base haplotypes differ collectively by 12 mutations from their
ancestral (see above) base haplotype, which translates 4050 ± 500 years to their ancestral
haplotype.
It should be noted that datasets of Indian R1a1 haplotypes are difficult to analyze, because they
typically represent a superposition of haplotypes from various sources, including those from the
ancient (pre-Indo-Aryan) ancestors, from the East European Plain, Central Asia, the Middle East,
etc. Since they all are present in various amounts and proportions, only analysis of their haplotype
trees can give meaningful results.
Other Scattered Ancient R1a1 Haplotypes in Asia
A "patchy" pattern of R1a1 was
created by the territorially blending of ancestors from the very ancient R1a1 (from more than 10,000
- 15,000 ybp) to the rather recent, the Aryan migrations. The tree in Figure 5 presents some
haplotypes from Nepal, which differ by 5 mutations from the East European Plain base haplotype,
pointing at a common ancestor of 7200 ybp. Some Indian haplotypes show 7-mutation difference from
the East European Plain haplotype with a common ancestor of 10,200 ybp. A Cambodian haplotype makes
9 mutations, which places a common ancestor of the East European Plain base haplotype and the
Cambodian haplotype at 14,000 ybp. Some haplotypes from Pakistan, Iran, Oman and Arab Emirates show
5 - 6 mutations, pointing to a common ancestor of 7000 - 9000 ybp. A group of ethnic minorities from
north-western China (Tu (aka Tu Fang "Tu aliens" of the Zhou stock,
the Scythians of the 17th c. BC), Xibe
(possibly, from "scattered", probably Türkic fragments joined with Tunguses at the turn of the
eras, politically a sedentary part of Kerulen Tatars, now indistinguishable from Tunguses),
Tatars (Tatars were an assembly of 3 autonomous groups of different origin
that included Hunnic, Uigur, and other Türkic refugees from different time periods),
Uigurs, Yugurs, Salars, Bonan and others) typically have their collective R1a1 common ancestor of
6900 ybp (Klyosov, 2010b). All of them reportedly have the following base haplotype, obtained from
the tree in Figure 7:
13 25 16 11 X X X 12 X 14 12 31
The value of DYS392 = 12 is suspect, because it is an unusual one for R1a1, including those from
Central Asia. However, almost all Asian haplotypes in (Shou et al., 2010) are reported as having
this "12" allele. The difference in 2.85 mutations with the base East European Plain or with the IE
Indian base haplotype (if DYS392 = 12 is correct) or 1.85 mutation (if DYS392 = 11 is correct)
places a common ancestor of the north-western Central Asian R1a1 haplotypes and the Russian/Indian
haplotypes to either 9350 ybp or 7925 ybp respectively. In any case, they are significantly more
ancient compared with the majority of the European haplotypes.
A Presumably Ancient (10,000 - 7700 ybp) R1a1 Population in Europe
There is a
distinct group of R1a1 bearers in Europe who have DYS392=13, while the overwhelming majority of R1a1
haplotypes in Europe and elsewhere has DYS392 = 11. The base haplotype of this group, coined "The
Old European branch" (Rozhanskii and Klyosov, 2009) is as follows:
13 25 15 11 13 14 12 12 10 14 13 31 — 16 9 10 10 11 25
14 19 31 12 15 15 15 — 10 11 19 23 16 16 17 17 36 38 11 11 — 11 8 17 17 8 12 10 8 12 10 12 22 22 15
10 12 12 13 8 13 23 22 12 12 11 13 11 11 12 12
This haplotype differs from the East European Plain R1a1 haplotype by 6 mutations in the first 12
markers, 12 mutations in the first 37 markers, 20 mutations in the first 37 markers, and 24
mutations in all the 67 markers, which places a common ancestor of their haplotypes at 6,800 ybp.
Besides, compared to the so-called North-western base (Atlantic modal
haplotype?) haplotype (Rozhanskii & Klyosov, 2009)
13 25 16 10 11 14 12 10 10 13 11 30 — 15 9 10 11 11 24
14 19 32 12 15 15 16 — 11 11 19 24 16 15 18 18 33 38 13 11 — 11 8 17 17 8 12 10 8 11 10 12 22 22 15
10 12 12 14 8 14 23 22 12 13 11 13 11 11 12 13
the difference is 9, 14, 24 and 29 mutations, respectively. Their common
ancestor lived ~7400 ybp.
Lastly, there is an additional comparison worthy of consideration, and it regards the Central
European branch (Rozhanskii & Klyosov, 2009) as follows:
13 25 16 10 11 14 12 12 11 13 11 29 — 16 9 10 11 11 23
14 20 32 12 15 15 16 — 11 11 19 23 17 16 18 19 34 38 14 11 — 11 8 17 17 8 11 10 8 12 10 12 21 22 15
10 12 12 13 8 14 25 21 13 12 11 13 11 11 12 13
Herein the difference is 9, 15, 25 and 34 mutations, respectively. Their common ancestor lived
~7700 ybp.
A series of 67 haplotypes of haplogroup R1a1 from the Balkans was published (Barac et al.,
2003a,b; Pericic et al.., 2005). In print, haplogroups were in 9-marker format, and the respective
haplotype tree is shown in Figure 10. Most of the tree contains typical European haplotypes
with a common ancestor of ~4500 ybp. However, the left branch is distinctive since it contains R1a1
haplotypes with DYS392 = 13, such as
12 24 16 10 12 15 X X X 13 13 29
12 24 15 11 12 15 X X X 13 13 29
13 24 14 11 11 11 X X X 13 13 29
This branch was calculated using both linear and the quadratic permutation methods. It obtained
.598 ± .071 mutations on average per marker, which resulted in 11,425 ± 1780 and 11,650 ± 1550 years
to a common ancestor, respectively (Klyosov, 2009a).
Calculations with short haplotypes are subject to high margins of error compared with extended 67
marker haplotypes. The authors of this study consider the timespans to a common ancestor of R1a1
haplotypes in Europe of 7400 - 7700 ybp to be more reliable in comparison to the 11,000 - 12,000
ybp. Haplotype databases continue to expand, and future studies will reveal the lower limit of "age"
of haplogroup R1a in Europe. Thus, dismissing the data of
Figure 10 would be premature.
Figure 10. The 9-marker 67-haplotype tree for the Balkans, haplogroup R1a1.
The tree was composed from data (Barac et al., 2003a, 2003b; Pericic et al., 2005) |
 |
9
Materials and Methods (ommitted)
Conclusion
The results of this study lend a support to the theory that haplogroup R1a arose in Central Asia,
apparently in South Siberia or the neighboring regions, such as Northern and/or Northwestern China,
around 20,000 years before present. The preceding history of the haplogroup is directly related to
the appearance of Europeoids (Caucasoids) ~58,000 ybp, likely in the vast triangle that stretched
from Western Europe through the East European Plain to the east and to Levant to the south, as it
was suggested in the preceding article (Klyosov & Rozhanskii, 2011). A subsequent sequence of SNP
mutations in Y chromosome, with the appearance of haplogroups NOP ~48,000 ybp and P ~ 38,000 ybp in
the course of their migration eastward to South Siberia eventually gave rise to haplogroup R ~30,000
ybp and R1 ~26,000 ybp, and then to haplogroup R1a and R1a1 ~20,000 ybp. The timeframe between the
appearance of R1a and R1a1 is uncertain.
At some point in time, the bearers of R1a1 began migration to the west, over Tibet and the
Himalayas, and not later than 12,000 ybp they were in the Hindustan. They continued their way across
the Iranian Plateau, along Anatolia and Asia Minor apparently between 10,000 and 9000 ybp. By 9000 -
8000 they arrived in the Balkans and spread westward over Europe and to the British Isles. At that
point, R1a1 still had DYS392 = 13 in their haplotypes, as did their brother haplogroup R1b1. This
marker is very slow, and mutates on average once in 3500 conditional generations. Somewhere on this
extended timescale, bearers of R1a1 (or the parent, upstream haplogroups) developed Proto-IE
language and carried it along during their journey from Central Asia to Europe. The earliest signs
of the language in Anatolia were detected by linguists, and dated by 9400 - 9600 - 10,100 ybp, which
coincides with the data of DNA genealogy that is described in this paper.
The arrival of R1a1 in Europe might be marked by known archaeological cultures in the Balkans and
Central/Eastern Europe, dated back to 9000 - 7000 ybp. Yet they also can be attributed to other
ancient haplogroups, such as I, J, E, G.
As the bearers of haplogroup R1b1a2 began to populate Europe after 4,800 ybp (the Bell Beakers
and other R1b1 migratory waves to Europe, including perhaps the Kurgan people, though their
identification and haplogroup assignment remains unclear), haplogroup R1a1 had moved to the East
European Plain around 4800 - 4600 ybp. From there R1a1 migrated (or moved as military expeditions)
to the south, east, and south-east as the historic Aryans. Dates for these movements are strikingly
similar, and they span 4200 and 3600 ybp. As a result, in Anatolia and Mitanni, South Ural, Iran,
India, and beyond the Ural Mountains, in South Siberia, in all those areas today’s linguists find
the same languages: the Aryan, or the Indo-European language, or the Iranian family of languages.
They all have the same Aryan roots. They founded common horse breeding terminology and shared
essentially the same vocabulary for household items, gods and religious terms, although sometimes
twisted due to "human factor" as found in India and Iran.
Currently, most of those with European R1a1 live in Eastern Europe, primarily in Russia (up to
62% of the population) and Poland, Ukraine, Belarus (up to 55% of the population in the last three
countries). In depth reports on their haplotype branches and distinct SNP (characteristic mutations
in the DNA) will be explored in forthcoming publications.
Acknowledgements
The authors are indebted to Laurie Sutherland for her valuable help with the preparation of the
manuscript.