Фраза, поставленная в заголовок, представляет довольно новую формулировку в науках о происхождении человека. До сих пор в популяционной генетике бытует мнение, что гаплогруппа R1a появилась где-то в южных степях, на территориях, где сейчас находятся современные Россия и Украина, по одним данным 15 тысяч лет назад, по другим 10 тыс лет назад, и переживала ледниковый период
в “украинском убежище”. Обе цифры вышли из-под руки Спенсера Уэллса без каких-либо пояснений, как без пояснений осталось и то, по какой причине он далее изменил 15 тыс лет на 10 тыс лет, и как это R1a могла переживать
“ледниковый период” 10 тыс лет назад на Украине, когда ледникового периода в те времена давно уже не было. Но популяционные генетики продолжают излагать и про ледниковый период, и про
“украинскую гаплогруппу” R1a, и про 15 тысяч лет назад. Никаких источников датировок никто из них не приводит, да и нет никаких этих расчетов ни у Уэллса, ни у других для
“южных степей”. Это – нормальное дело для популяционной генетики.
В то же время немало данных указывают на Южную Сибирь и на её алтайский регион как место обитания древних европеоидов. Это было показано как старыми работами по анализу
“генетических расстояний” в полиморфных формах белков (серия работ А.Ф. Назаровой 1980-х и 1990-х годов), так и исследованиями по переходу носителей гаплогруппы Q из Южной Сибири, и, видимо, именно из Алтайского региона через Берингов мост в Америку. Поскольку прибытие носителей гаплогруппы Q в Америку датируется по разным данным от 20 до 12 тысяч лет назад, то они должны были покинуть Южную Сибирь не позднее 40-30 тысяч лет назад (обычно скорость древних миграций принимается за 1 км в год).
По расчетам, гаплогруппа Q образовалась из сводной гаплогруппы Р в интервале 45-40 тысяч лет назад. Тогда гаплогруппа Р уже должна была быть в Южной Сибири в те времена, и позднее образовать гаплогруппы R, R1, R1a и R1b. Все эти гаплогруппы – европеоидные по антропологии, и европеоидными они и пришли в Европу – R1a примерно 11-9 тысяч лет назад на Балканы, R1b – около 10 тыс лет назад на Русскую равнину, и около 5 тысяч лет назад в западную и центральную Европу (а также на Балканы, в юго-восточную Европу).
1511
Вот такой, казалось бы, парадокс – европеоиды получили свое название от Европы, в которую прибыли после долгих и далеких миграций из Центральной Азии. Но на самом деле все правильно, если принять возможность того, что европеоиды в самом деле образовались в Европе в виде сводной гаплогруппы FT, из которой выделились две ветви – IJK осталась в Европе, а NO+P ушли на восток, в Южную Сибирь, где гаплогруппа Р и осела на долгие 20 тысяч лет, перед своим (в потомках гаплогрупп R1a и R1b) началом возвращения в Европу.
Такие длинные миграции традиционно воспринимаются с недоверием в академических кругах исследователей истории и лингвистики. У них просто нет методологического аппарата для изучения столь длинных миграций – на многие тысячи километров и на протяжении многих тысяч лет, порой десятков тысяч лет. Обычно сходство материальных признаков на разных концах континентов, а то и на разных континентах принимается за случайные совпадения. Я не раз читал в качестве подобных аргументов то, что поскольку знак свастики, например, найден у древних индейцев, где ариев и быть не должно, значит, случайное совпадение, одинаковое представление солярного знака. Меня этот аргумент, признаться, никогда не убеждал, но отвести его - не было объективных данных. Сейчас они появились, в виде практически одинаковой керамики с десятками одинаковых признаков у европеоидов (maybe Europeans?) (и ариев) и у американских индейцев (частное сообщение Е. Мироновой), но это предмет другого разговора.
ДНК-генеалогия приводит новые и неопровержимые доказательства таких длинных миграций. Гаплогруппа R1a была найдена в северном Китае, где в ряде местных народностей и этнических групп ее доля достигает 30%. И главное даже не это, а то, что их гаплотипы настолько испещрены мутациями, как редкие (или вообще никакие) гаплотипы других гаплогрупп. Гаплотипы группы R1a в северном Китае доступны только короткие, но тем и характерны, потому что короткие гаплотипы обычно намного более стабильны, чем протяженные. Точность датировок по ним меньше, но при таких
“глобальных” мутациях точность уже не столь важна, важен сам феномен. Северо-китайские гаплотипы дали датировку 20 тысяч лет до общего предка гаплогруппы R1a в том регионе.
1512
Базовый гаплотип их 5-маркерных гаплотипов
13 Х 14 Х Х Х Х 12 Х 13 Х 30
имел только одно характерное отличие от гаплотипов R1a Русской равнины – в третьем маркере (DYS19) у последних обычно 16 или 17, китайских – 14.
Алтайские гаплотипы R1a были исследованы нами в работе (Klyosov and Rozhanskii, 2012), и найдено, что их общий предок жил совсем недавно по историческим меркам – всего 825±320 лет назад, и его базовый гаплотип был
13 26 16 11 11 17 Х 12 11 14 Х 31
Он заметно отличается от базового гаплотипа R1a Русской равнины парой DYS385=11-17, которая на Русской равнине равна 11-14
13 26 16 11 11 14 12 12 10 13 11 30
Общий предок этих двух базовых гаплотипов жил 8100 лет назад (Klyosov and Rozhanskii, 2012). В целом перекрестное сопоставление базовых гаплотипов разных алтайских популяций привело к расчетам времен до общих предков европейских и алтайских гаплотипов группы R1a в интервале 10,400 и 7,300 лет назад (ссылка там же). Та же картина была получена и для тувинских гаплотипов, в целом, общий предок европейских, алтайских и тувинских гаплотипов жил 10,000-10,400 лет назад (Klyosov and Rozhanskii, 2012).
Здесь следует отметить, что рассмотрениям популяционных генетиков присуща одна и та же ошибка, которая постоянно повторяется в их
“академических” публикациях. А именно, статическая картина и отсутствие исторического понимания. Если они видят гаплотип группы R1a в Азии, так он непременно
“пришел из Европы”, его тут же называют “скифским”, “курганной культуры”, “индоевропейским”, не имея к этому ни малейшего основания. Иначе говоря, идут
“рассуждения по понятиям”, а не по науке. Не по науке – потому что датировки они считать не умеют и в рассмотрения-рассуждения их не включают.
На самом деле данные и расчеты показывают, что в алтайском регионе, Китае, Монголии, Южной Сибири, Центральной Азии (эти регионы пересекаются), есть по меньшей мере две группы гаплотипов группы R1a.
Одна группа – исходная, древнейшая в отношении их общих предков, не европейская и не “индоевропейская”, не арийская, а сугубо автохтонная.
Эта группа фрагментирована, прошла различные бутылочные горлышки популяций, и представляет сейчас потомков предковых линий давностью от 600-800 лет назад до 7 тысяч лет назад
(Mesolithic), в редких случаях – до 20 тысяч лет назад
(Upper Paleolithic), как у серии северо-китайских гаплотипов. Гаплотипы у этих серий заметно отличаются от гаплотипов Русской равнины.
1513
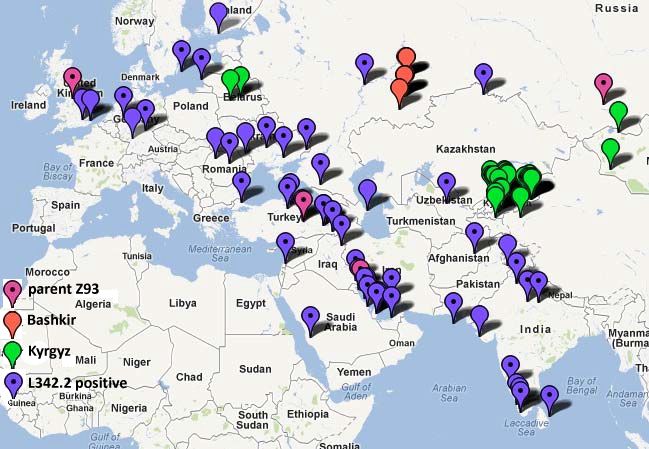
Другая группа – это гаплотипы, действительно “пришедшие” из Европы 4000-3000 лет назад, со своими, как правило, арийскими,
“индоевропейскими” носителями группы R1a. Если они типируются на субклады R1a, оно скорее всего покажут юго-восточный субклад L342.2 (R1a1a1h1-L342.2, Middle East, India, eastern part of the E.European Plain, hence South-Eastern
branch) и его производные.
Map 3 (Klyosov and Rozhanskii, 2012)
Map of South-Eastern branch, with birthplaces of furthermost ancestors recorded in available
databases.
Magenta pins denote a parent Z93 branch
red and green pins correspond to Bashkir and Kyrgyz branches, respectively. The rest of the branch
(L342.2 positive) is deep-blue.
The most represented Ashkenazy Jewish branch (not shown) covers densely an area of historical
Polish-Lithuanian Commonwealth.
Поэтому мумии Таримского бассейна, например, с датировкой 4000 лет назад, могут быть или автохтонные, или
“индоевропейские”, и не стоит торопиться называть их “европейцами”, “скифами”, “курганниками”. К этому, как было сказано, нет никаких оснований. Здесь помогли бы гаплотипы, но они, к сожалению, или не определялись, или не публиковались. Аргументы, что
“на них была одежда типа шотландского пледа” тоже не являются состоятельными, потому что для состоятельности нужно показать, что этот тип одежды не был, напротив, принесен с Алтая в Европу. Рассмотрение подобных альтернатив не присуще популяционным генетикам.
Решению этого вопроса пока не помогает и археология. Рассмотрение афанасьевской культуры могло бы, наверное, внести ясность в решение проблемы, хотя бы частично, но оказалось, что в афанасьевской культуре практически отсутствуют объективные датировки, и откуда прибыли ее люди – остается неизученным, то ли они автохтонные, то ли
“европейцы”. Анализ ископаемых костных остатков не предмет гаплогрупп не проводился.
Обычное недоумение о связи происхождения гаплогрупп R1a, по сути европеоидной, с Алтаем, сводится к тому, что носители R1a на Алтае (и в прилегающих регионах) сейчас – типичные монголоиды и говорят на языках тюркской группы. Вопрос, который обычно задается – какие же это
“индоевропейцы”, когда это тюрки-монголоиды? На это можно опять привести комментарий выше об отсутствии исторического видения, типа
“что вижу, о том и пою”. Гаплогруппа R1a на Алтае, как сообщалось выше, прошла бутылочное горлышко популяции всего 800-900 лет назад. Что это означает? То, что это уже не те европеоидные предки, которые там жили тысячелетия назад, хотя гаплогруппа сохранилась, хотя бы частично. Это означает, что ранее 800- 900 лет назад (или в те времена) эта популяция была почти истреблена – либо в результате природных катаклизмов, либо – скорее всего – врагами.
Выжившие носители R1a и их потомки по своему генотипу, а, следовательно, фенотипу, были уже другими людьми. То, что они перешли на тюркский язык, показывает, кто могли быть их недругами. Хотя причиной могли быть и тюркоязычные подруги и жены. Если недруги - по времени это могли быть китайские или монгольские нашествия 11-13 вв нашей эры. Гаплогруппа R1a у многих потомков осталась, а значит, уцелела у (немногих) выживших предков, но жены были уже не европеоидные, а монголоидные.
|
We know of well-documented historical events that illuminate the demographical changes leading to
and in the early 1st mill. AD. These are consecutive relocations of the Türkic nomadic states of
Oguzes (740), Kangars/Bechens (colloquial Pechenegs, 750), Karluks (colloquial Karakhanids, 920),
Kimaks/Kipchaks (colloquial Polovetses, 990) from the greater Altai area to the west. Assuming that
2/3 of the population have relocated, 1/3 of the population minus military and hardship losses
stayed put, and carried on their genotype. The new demographic status at the destinations was a blend of the previous
population and the newcomers from the immediate east, linguistically also Türkic, but with
relatively higher proportion of the eastern Eurasian Y-DNA and mtDNA genes. The new geographical layout remained relatively stable for 200 years, until the turmoil of the
Chingizkhan. Chingizkhan subjugated and wiped out the surrounding linguistically Türkic and Mongolic
people, but his wipeout achievement zeroed on the Mongolic division of the three-division Tatars,
leaving the Uigur division intact, and also leaving intact the nominally Tatar component of the
Kimak Kaganate. Chingizkhan was a recognized offspring of the Uigur Kaganate dynastic line Jalair,
via a Borjigin ancestor, and supposedly belonged to the Uigur Jalair Y-DNA line. As a result, after
the “Tatar wipe-out", the subsequent movement headed by few thousand Mongols became known as a Tatar
movement.
The Tatar movement, demographically nearly exclusively linguistically Türkic, brought about
further demographic outflows from the greater Altai area. Assuming that 2/3 of the population have
relocated westward, the 1/3 of the population minus military and hardship losses stayed put, and
carried on their genotype.
That is “the enemies - timewise they could be Chinese or Mongolian invasions of the 11-13 cc.
AD". There were no Chinese or Mongolian invasions of the 11-13 cc. AD, the Chinese were cornered and
could not fend for themselves, and Mongolian nomadic drifters were just filling in the voids left by
the “Tatar" migrations. On the other end of the Urals, at that period occurred a corresponding
demographic change with the opposite effect, in the Eastern Europe formed and endured a new numerous
nation that absorbed predominantly Türkic existing populations and the new Türkic migrants from the
east, known as Kipchak Khanate and Tatar Khanate (colloquial Golden Horde, 1240).
Demographically, few of the continuous 6-centuries long Altai migrations to the Eastern Europe
reverted back to their greater Altai domicile. The majority were absorbed by the new powers-to-be,
adding their genetic markers to the local ethnicities. It is inconceivable that the genetic markers
of the early Altaians would not be found in the Eastern Europe and in the Balkans; the genetic
markers remaining in situ in the C.Asia had to be subjected to the stresses and dilutions of the coming centuries.
By the early 13th c., of the mid-1st mill. AD population, in the Altai probably remained only 10%
(1/32) of the early Altaian population, 90% of the early Altaians having moved to the
Middle Asia and Eastern Europe. At least partially, the outflow was replenished by the kindred genotypes migrating from the
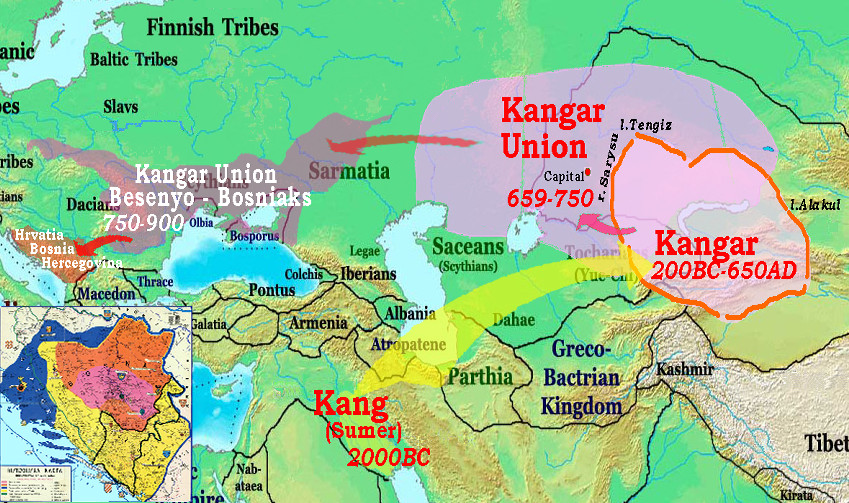
east. And then in 1650 fell a Jungar genocide. Numerous populations can be traced individually, for example the Kangar anabasis below:
The influence of the female Mongoloid genotype can't be underestimated, and should not be
overestimated. The Mongoloid admixture, also termed Lapanoid and Uralic, is traced in the eastern and central
European population from the Paleolithic times; every study of the Kurgan people and steppe nomads from the later Mesolithic times to the
Middle Ages noted their Caucasoid/Mongoloid admixture, and notoriously more notable Mongoloid appearance of the
females than of the males. To suggest a supposed linguistic replacement to the effects of the
Türkic-speaking mistresses and wives mass-deranging the noble IE speakers is an open-ended
proposition that affects all ages down to the Paleolithic times.
For horse-mounted nomads, the greatest influential demographical factors are migration (exodus),
followed by genocides, followed by exogamic law. |
1514
Именно это изменило и генотип, и фенотип потомков при сохранении мужской хромосомы и ее гаплогруппы R1a. Можно ожидать, следовательно, сдвига спектра мтДНК в сторону азиатских, монголоидных вариантов митохондриальных гаплогрупп. Как будет видно ниже, именно так и оказалось.
Понятно, что таком состоянии вопроса новые научные публикации, на новом материале и с обоснованными выводами представляют несомненные интерес и важность. И вот в этом, 2012 году, появились две публикации на
“алтайскую тему”. Первая – совместная статья специалистов из Пенсильванского униветситета (Matthew Dulik, Theodore Schurr и др.), Института цитологии и генетики Сибирского отделения РАН (Сергей Жаданов и Людмила Осипова) и Института общей генетики и цитологии Казахстана (Айкен Аскапули), под названием
“Вариации мтДНК и Y хромосомы свидетельствуют о недавнем общем предке америндов и автохтонных алтайцев”. Вторая – статья испанских авторов (Барселона) Gonzales-Ruiz, Malgosa и др.
“Происхождение смешивания популяций востока и запада алтайского региона (Центральная Азия)”.
Надежды на появление продвинутых работ, как водится, оправдались только частично. Обе работы содержат интересные находки, которые, правда, в значительной мере тонут в болоте несообразностей. Более содержательной оказалась первая работа, но она на наши вопросы, изложенные выше, не ответила. Тем не менее, дала сырой материал, на основании которого можно было провести некоторые (но вполне содержательные) расчеты, показанные ниже.
Начнем со второй, значительно более простенькой (в том контексте, который изложен выше) работы.
Статья Gonzalez-Ruiz et al. (2012)
Gonzales-Ruiz и соавторы исследовали мтДНК в костных остатках Пазырыкской культуры. Это – археологическая культура железного века (3- 5 вв до н.э. , хотя некоторые опускают дату до 6-го века до н.э.)
(see Marsadolov L.S., 1994, Pazyryk Timing, and
A.Yu.Alekseev et. al., 2001, Chronology of Eurasian Scythian Antiquities
Born by New Archaeological and 14C Data), которую
(archeologically) относят к “восточному скифскому кругу”. Регион – горный Алтай и смежные территории Алтая, Казахстана и Монголии. Основное занятие – кочевое скотоводство. Высказывается предположение, что пазырыкская культура – производная от афанасьевской. Ее основные антропологические типы - долихокранный европеоидный с высоким и широким лицом (видимо, носители гаплогруппы R1a), брахикранный монголоидный с невысоким лицом и мезодолихокранный монголоидный с высоким лицом (последние могут быть носителями гаплогрупп С, N, Q, и парным им мтДНК).
1515
Надо сказать, что предыдущий параграф в статье почти не изложен. Вместо этого авторы много говорят о том, что Алтай – граница западных и восточных евразийских степей, о особенно много
о “скифах”, которые были “европеоидны”
("Caucasoid") и говорили на “индоевропейских языках”. В общем, стандартный букет популяционно-генетических установок, включая упоминания Геродота. Никаких других данных, что изучаемые авторами костные остатки принадлежали именно скифам, в статье нет, и неясно, зачем о скифах нужно было вообще писать. Вообще неизвестно, скифы ли это. Тут же и про
“номадический пасторализм”, хотя кости про это тоже ничего не говорят. Короче, мощным фоном идет то, что к содержанию материала никакого отношения не имеет
(probably, the background summarizes contents and terminology of the
bibliography). Но так принято в популяционной генетике. То, что в захоронениях нашли европеоидные антропологические особенности, тут же подается, как
“скифы имели европейскую морфологию”. То, что европеоидность могла остаться от автохтонных европеоидов, авторам в голову не пришло. Популяционная генетика.
Авторы изучали мтДНК (HVRI, гипервариабельная область) 19-ти скелетных остатков монгольского Алтая из четырех захоронений, относящихся к бронзовому (три мтДНК)
(3000-700BC) и железному (16 мтДНК)
(800-300BC) веку.
|
Studies of pre-historical periods without reasonably accurate dating is senseless, and any
attributions and conclusions are incredible. |
Было постулировано (точнее, принято как данность) что гаплогруппы мтДНК подразделяются на три региональных происхождения:
-- Западно-евразийские: R0 - R0a’b, HV, N1, JT, UK, W, X
-- Восточно-евразийские: М – C, D, G, Z, M9, M10, M11, M13, A, B, F, N9a
-- Южно-азиатские: M*, U1a-c, U9, R*, R1-R2, R5-R6, N1d.
Дальше пошли типичные попгенетические корреляции, типа “разнообразия генов”, “разнообразия нуклеотидов”, Арлекин, FST,
“парные генетические расстояния между популяциями” и прочее, чему смысла в ДНК-генеалогии нет, но есть какой-то тайный смысл в популяционной генетике, что и хорошо. Чем излагать эту алхимию, сразу посмотрим, что это по сути дало. Это будет конструктивнее.
Итак, из 19 ископаемых гаплогрупп 11 оказались A, C, D и G, то есть “восточно-евразийские”, 9 оказались J, K, HV, U и T, то есть
“западно- евразийскими”. Авторы предпочли эти цифры – 9 и 11 – напрямую не давать, и указать “58%” и
“42%”, иначе уже слишком малы эти количества.
На самом деле из первых 11 гаплогрупп 7 оказались гаплогруппы D – все три гаплогруппы “бронзового века” и еще четыре – железного века. По одной были А, С и G, все из железного века.
1516
Еще в железном веке в алтайской выборке были найдены три К, два U (U5a1), по одной J, HV, и Т. Это позволило авторам сделать вывод, что в бронзовом веке все (три) гаплогруппы были
“восточные”, а в железном веке к ним добавились разнообразные “западные”. Это и есть “admixture”. В общем, может быть. Правда, из бронзового века были всего три образца, а из железного – 16. Интересно, какое было бы распределение, если бы было наоборот, 16 из бронзового, и 3 из железного? Оно, явно, было бы другим. Тем не менее, авторы назвали пазырыкскую популяцию
“генетически гомогенной”.
Если принимать картину как данность, то получается, что невестами и женами алтайцев (в основном R1a, как будет видно ниже) были восточно- азиатские женщины (наверное, монголоидные), а в железном веке добавились западноевразийские женщины. Либо сами приходили, либо их умыкали.
Вот, собственно, и вся статья. Все остальное – общие разговоры. Много из той статьи, конечно, не почерпнуть. Хотя сам факт анализа ископаемых мтДНК – уже немалое достижение, требующее определенной технической научной школы.
Cтатья Dulik et al. (2012)
Статья Dulik et al., как было упомянуто, значительно более продвинутая.
Правда, основной вопрос, который она рассматривает, а именно о происхождении америндов, и приходит к выводу, что они из Южной Сибири, давно решен в литературе. И то, что общими между ними являются гаплогруппы Q и C, тоже давно решен. Авторы углубляются в детали, какие именно субклады являются у них общими, но это, на мой взгляд, мало что дало, кроме общих упражнений на эту тему. Ну, нашли общие субклады (на самом деле больше не нашли, чем нашли), и что это даст, на какую загадку ответит? Более того, авторы запутались между
“генеалогической” и “популяционной” скоростями мутаций, и в итоге пришли к категорически неверному выводу, совершенно смазав дискуссию и выводы.
Огорчительно, что авторы оказались совершенно не знакомы с современной литературой на тему скоростей мутаций. Например, автор этих строк опубликовал еще три года назад обстоятельные статьи в журналах
J. Genet. Geneal. (2009) и Human Genetics (2009) о том, какие на самом деле величины констант скоростей мутаций и как они калибруются.
1517
Ну, не читают авторы литературу по скоростям мутаций, тогда зачем публиковать на эту тему статьи, увы, безграмотные? Это уж не говоря о том, что многочисленные форумы по косточкам уже давно разнесли
“популяционные скорости Животовского”, а авторы об этом, видимо, и не догадываются. Это как же надо не уважать предмет своих исследований, настолько им не интересоваться, чтобы не читать форумы, наконец, не читать журнал
“Вестник Академии ДНК-генеалогии”, в полусотне выпусков которого – с 2008 года – эти константы скорости мутаций разбирались десятки раз, в том числе многократно было показано, что
“популяционные скорости Животовского” - это для реальных систем сущая бессмыслица, более того, по сути это продукт подгонки и откровенной манипуляции данными. Об этом были отдельные статьи в Вестнике. И тем не менее, авторы ссылаются на Животовского, хотя, похоже, уже понимают, что что-то там не так. И потому считают параллельно
и “генеалогически”, и “эволюционно”, что приводит вообще к полной каше в результатах расчетов. Так и пишут, что если, мол, считать
“генеалогически”, то получается недавно, а если “эволюционно”, то бронзовый век. Ну нельзя же так. Хотя, впрочем,
“популяционная генетика”, там все можно. Как и в шаманстве.
Но самое забавное даже не это. Авторы в итоге пришли в тому, что “популяционная скорость” более правильно описывает данные, лучше стыкуясь с данными истории. Так,
“генеалогические” скорости для южно- американских Q1a3a1a* дают времена жизни общих предков (по данным авторов) в Америке 4.9 тыс лет и 7.7 тыс лет,
а “по Животовскому” 22.0 и 13.4 тыс лет, значит, вторые правильнее.
Это, конечно, уму непостижимо. Слышали ли авторы про “бутылочные горлышки популяции”? Откуда они вдруг решили, что любая популяция при расчетах дает именно
“истинное” значение времени жизни древнейшего общего предка? Например, по данным самих же авторов серия алтайских Q1a3a* (которая должна быть старше по филогении, чем Q1a3a1a*) на 19-ти 17-маркерных гаплотипов дает всего 29 мутаций, от базового гаплотипа
13 24 13 10 15
18 Х Х 13 14
14 17 –
16 14 19 11 16
11 22
причем в отмеченных маркерах вообще нет мутаций. Ясно, что общий предок этих 19 гаплотипов – недавний. Действительно, 29/19/0.034 = 45
→ 47 условных поколений (25 лет в каждом), то есть 1175±250 лет до общего предка. А должно быть, по
“логике” авторов, не менее 20 тысяч лет. Значит, “метод неправильный”, не так ли? А это просто предки в потомках не выжили, ДНК-генеалогическая линия почти пресеклась, популяция прошла бутылочное горлышко, или
“генетический дрейф”, как любят говорить попгенетики. Короче, недавно жил общий предок данной серии, и ничего неправильного здесь нет. Так и в Америке для серии Q1a3a1a* гаплотипов. Не надо ее искуственно задирать в три раза с помощью ущербного метода.
1518
Не в том, конечно, “продвинутость” статьи, что неправильно считали, и что “открывали Америку”, которая давно открыта. А в том, что поместили в статью хорошую подборку гаплотипов разных алтайских популяций, целых 189 гаплотипов в 17-маркерном формате. Лучше бы гаплотипы подлиннее, но и на том хорошо. Лучше бы субклады у R1a1a1 выявили, но и на том хорошо. А то измерили только R1a1a-M17 (R1a1a) и R1a1a1-M417 (R1a1a1) [первый снип можно было бы и не определять, он в данной ситуации лишний, поскольку все имели М417], и на этом остановились, неверно отметив R1a1a1* (со звездочкой), как будто у него нет нисходящих снипов. То, что у него не оказалось M458 (центрально-европейская и западно- славянская ветви) и L365 (северо-европейская ветвь), так их и не могло быть. Известно, что на востоке их нет, кроме недавних
“туристов”. А вот измерить L342.2 было бы в самый раз, он-то и разделил бы автохтонные субклады и “индоевропейские”. Жаль, авторы об этом снипе, наверное, и не знали.
Авторы нашли, что изученные популяции имели в основном “восточно-евразийские мтДНК гаплогруппы”, что целом противоречит предыдущей, цитированной выше работе, согласно которой начиная с железного века началось активное
“смешивание” мтДНК. В действительности же, гаплогрупп A, C, D и G оказалось большинство у всех тестированных популяций:
-- у челканов (Chelkandy, N.Altai, a part of Tubalars) (38%), еще 19% N9a, остальные минорные;
-- у кумандинов (Kumandy, Kumans, N.Altai. with seoks Chabat, So/Solu) (62%, остальные все минорные);
-- у тубаларов (N.Altai, Turgeshes with seoks Chagat/Chygat of Tele stock,
Chalkanyg; Kumysh/Komlyash/Komnosh aka Black Tatars, Mountain/ Taiga Tatars, Kumysh = Silver; Kuzen,
Palan-Komdosh/Bolan, Tiber, Togus/Tokuz, Turgesh/Kergesh, Yalan, Yus, Yus-Shanmai) (64% и 53% у двух популяций, остальные все минорные);
-- у шоров (N.Altai, with seoks Aba/Aban, Chelei/Teleut, Kalar, Chelei/Elei,
total 22 seoks) (50%), еще 14% у F1, остальные минорные;
-- у алтай-кижи (N.Altai, with 36 seoks Irkit, Naiman, Kypchak, Mundus
the largest seoks; Tele 30%, Türks/Ashina Türks 22%, Enisei Kyrgyzes 12%, Naimans 10%) (48% и 36% у двух популяций, остальные все минорные),
-- у теленгитов (S.Altai, Tele tribes Telengut, Telengit, and Tele) (40%), еще 15% гаплогруппы В, остальные все минорные, и
-- у телеутов (S.Altai, Tele tribes Telengut, Telengit, and Tele) (51%, остальные все минорные).
Это важно, и подтверждает высказанную выше гипотезу, что несмотря на европеоидную
(Y-DNA) гаплогруппу R1a, монголоидные женщины превратили носителей R1a из европеоидов в монголоидов, да еще и сменили их язык на тюркский.
|
In retrospective study of the Türkic tribes' origin, Altai served as a refuge, over millenniums
refugees from many surrounding lands found safety in Altai. The layers of refugees superimposed,
intermixed, and merged like layers of old paint on an antique furniture. Peeling one layer after
another, in many cases the origin of the Altai tribes can be tracked down to their first naming in
the Chinese dynastic chronicles, in the works of antique Greek authors, and other antique sources.
For the Türkic tribes, Altai, like just like Caucasus and Khingan, was a shelter with ongoing
historical merging, and not a source from which emanated numerous Türkic tribes and peoples.
Complimentary to Altai, the Minusinsk depression of the “Karasuk time" in the middle of the 2nd
millennium - 7th century BC was a stopover location in the spread of some Türkic tribes, possibly
together with the Ket tribes. A good case study example is the case of Tokuz-Oguz (Nine tribe
confederation, aka High Wheels, Ch. Dinlin/Dingling) tribes Apa/Atie/Adie, Beiho/Huiho/Husie or
Uigurs, Dubo/Tubo, Guligan/Kurykan = Sakha/Yakut, Kibi/Kipi, Paegu/Baegu, Pugu, Seyanto, and Tunlo,
which in the words of Chinese annals constituted a “special generation of Tele", and the tribes Fulo/Buri,
Fuliku/Buriku, Hun, Husie/Husa, Sekis (Se/Saka), Taki, Teleer, and Telengit/Tolanko/Dolange, which
also were treated as Tele tribes.
A good case study example is the case of Tokuz-Oguz (Nine tribe confederation, aka High Weels,
Ch. Dinlin/Dingling) tribes Apa/Atie/Adie, Beiho/Huiho/Husie or Uigurs, Dubo/Tubo, Guligan/Kurykan =
Sakha/Yakut, Kibi/Kipi, Paegu/Baegu, Pugu, Seyanto, and Tunlo, which in the words of Chinese annals
constituted a “special generation of Tele", and the tribes Fulo/Buri, Fuliku/Buriku, Hun, Husie/Husa,
Sekis (Se/Saka), Taki, Teleer, and Telengit/Tolanko/Dolange, which also were treated as Tele tribes.
It has long been established that Tele tribes were Türkic-speaking. A corollary of that is that
the mixed phenotypically Caucasoid/Mongoloid men with genetically Caucasoid/Mongoloid genes, and the
mixed phenotypically Caucasoid/Mongoloid women with genetically Caucasoid/Mongoloid genes spoke the
Türkic language. What was the language of the female's ancestors, as well as the male contribution
from the eastern lineages, may be clarified one day with the help of the ongoing genetic studies,
but a linguistic mass-conversion of the noble IE speakers, without a grain of detected noble IE
substrate in the body of the 42 mongrel Türkic languages, is a pathetic pipe dream.
Not all of the Tele tribes were nomadic horsebreeders. Not all of the Tele tribes were elite
tribes either. The Chinese annals illuminated the useful, visible, and dangerous, leaving in the
shade non-prominent and dependent kyshtym tribes (kyshtym = allied dependency/tributary). These
tribes economically were at a different level from the horsebreeders, their traditional sedentary
subsistence was in hunting, fishing, farming, small ungulate husbandry, charcoal, metal smelting,
metalwork, and such. At the turn of the eras, numerous Tele tribes and confederations occupied
sub-taiga woodland belt from Khingai in the east to the Urals in the west. Neither Chinese annals,
nor the later writers reported on social relations within the Türkic confederations, but the first
records on the Türkic tribes that fled to Altai in the 16th-17th cc. reported that the elite tribes
were fleeing with their kyshtyms, and the local elite Enisei Kirgiz tribe had numerous kyshtym
dependents across Altai and sub-taiga woodland belt. Understanding the ethnic and social
relations is a necessary condition for genetic sampling and conclusions of the ancient and modern
Altaians.
One of the Tele tribes was Shibir, it was an elite tribe that gave the Türkic Kaganate at least
one Kagan (609-619), probably from the Ashina Kagan clan and Sibir Hatun clan. Shibir tribe is
listed among the Tele tribes that Juchi subdued in 1207, along with the forest tribes of Kesdiin,
Bait, Tuhas, Tenlek, Toeles, Tas, and Bachjigi. Shibir was the westernmost tribe, and the
easternmost tribe Bachjigi was occupying the Sayan-Altai uplands. At about the same time, the tribe
Sabir/Savir/Subar/Suvar/Suvar gained an upper hand and a title “King" of the Armenian chroniclers
over the Kayi and Masgut tribes in the E.Caspian seaboard in the Caucasus. The first records on the
nomadic Sabirs come from the Babylonian cuneiform tablets of 23rd-22nd cc. BC, along with the
Mesopotamian horse nomadic tribes Kuman, Quti, Lulu, and Turuk. Thus, we have literary traces on the
tribe Sabir from the 23rd-22nd cc. BC to modernity, from the Mesopotamia to Khingan, illustrating
the author's point on popgenetic's static picture and a lack of historical comprehension. |
1519
Переходим к самому важному для нас (в ДНК-генеалогии) – гаплотипам. Из 189 гаплотипов 75, то есть 40%, было R1a, из них большинство – 60 гаплотипов R1a - у популяции алтай-кижи
(Tele 30%, Türks/Ashina Türks 22%, Enisei Kyrgyzes 12%, Naimans 10%). Для сравнения – R1b гаплотипов было всего 6 единиц, или 3%, все в популяции кумандинов. На втором месте по численности была гаплогруппа Q, cуммарно 34 гаплотипа (18%), в пяти субкладах. Гаплогруппа С (в двух субкладах, C3* и С3с1) включала 24 гаплотипа (13%). Гаплогруппа N в четырех субкладах (N1*, N1b*, N1c* и N1c1 [последнего всего два гаплотипа]) включала 22 гаплотипа (12%). Остальные гаплогруппы – D3a (6 гаплотипов), J2a (3 гаплотипа), E1b1b1c, I2a и L (по одному гаплотипу). Это все, так сказать, опять популяционная генетика, и относится к тому, что есть СЕЙЧАС. К ДНК-генеалогии это не имеет практически никакого отношения. Все эти единичные гаплотипы из 189, как правило, случайны, например, из групп I2a, E1b, J2а в Южной Сибири. Что не случайно – это 75 гаплотипов европеоидной (по происхождению) гаплогруппы R1a, причем если на западе они имеют в основном мтДНК группы Н, то на Алтае – в основном мтДНК группы С. Вот и причина их монголоидности.
Altai-kiji (Tele 30%, Türks/Ashina Türks 22%, Enisei Kyrgyzes
12%)
To put things in perspective, here is an example of 1897 census, totaling 19,000 Altai-kiji people.
The compatible number for 2002 census was 62,000. The rate of population growth points to significant dilution
of the 1897 genetic inventory. But the number of 1897 census, taken by taxing district (volost, a distorted Türkic ulus with a prosthetic v) with mixed
population, was already extremely mixed, with the Altai-kiji proper constituting only 8% (1520 total)
of the listed Altai-kiji. A blind sampling of the modern Altai-kiji is ethnically as
informative as testing for its origins a herring gumbo soup (data from
L.P.Potapov, 1969, “Ethnic composition and origin of Altaians”, Science, Leningrad).
| 1 |
Todosh |
2978 persons |
19 |
Merkut |
166 persons |
| 2 |
Irkit |
1928 |
20 |
Koojo |
139 |
| 3 |
Naiman |
1912 |
21 |
Kaan |
118 |
| 4 |
Kypchak |
1681 |
22 |
Almat |
105 |
| 5 |
Mundus |
1342 |
23 |
Chagandyk |
89 |
| 6 |
Teles |
1333 |
24 |
Modor |
72 |
| 7 |
Kergil (Turgesh) |
1035 |
25 |
Tumat |
72 |
| 8 |
Tongjoan |
974 |
26 |
Derbet |
56 |
| 9 |
Soen |
684 |
27 |
Bogus-han |
49 |
| 10 |
Kobek |
565 |
28 |
Yaryk |
28 |
| 11 |
Chajats |
558 |
29 |
Yus |
19 |
| 12 |
Ochy |
528 |
30 |
Saal |
15 |
| 13 |
Bailagas |
398 |
31 |
Purut |
9 |
| 14 |
Koobaly |
348 |
32 |
Elik |
7 |
| 15 |
Dieti-Sary |
295 |
33 |
Mogol |
3 |
| 16 |
Olun |
275 |
34 |
Meret |
2 |
| 17 |
(garbled) |
245 |
35 |
Togus |
1 |
| 18 |
Gandi |
229 |
36 |
Suuzar |
1 |
| Total |
19,000 |
|
|
|
|
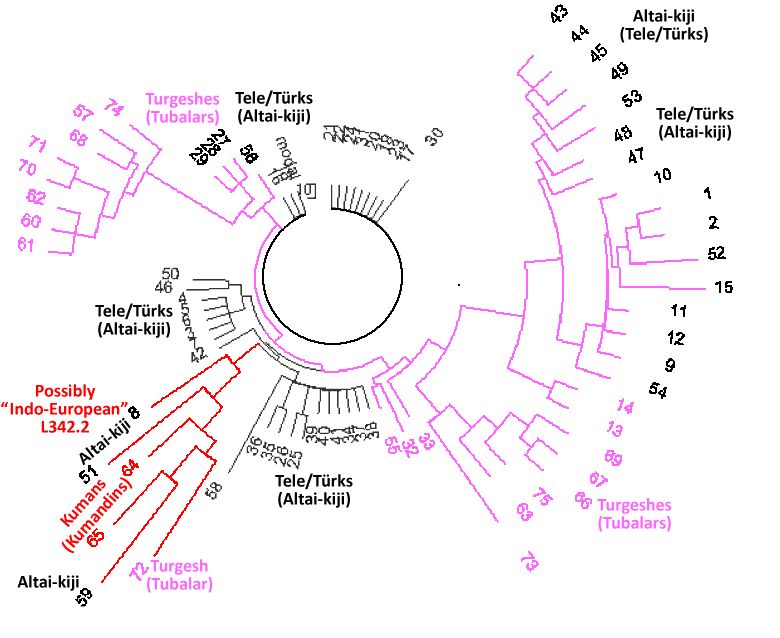
Рис. 1. Дерево из 75 гаплотипов гаплогруппы R1a1a1-M417 на Алтае.
Верхняя левая ветвь – тубалары (57, 68, 70, 71, 74), челканы (60, 61, 62) и алтай-кижи (57). Нижняя правая ветвь – подобный состав: тубалары (66, 67, 69, 73, 75) и один челкан (63). Нижняя левая ветвь – смешанная: кумандины (64, 65), тубалар (72), алтай-кижи (8, 51, 59). Остальные гаплотипы – алтай-кижи.
(color coding added, ethnic affiliation conditional to statistical
probability of fraction in population and randomness of sampling)
1520
На рис.1 приведено дерево 75 гаплотипов гаплогруппы R1a на Алтае.
Видно, что дерево состоит из нескольких совершенно различных ветвей. Столь характерное дерево редко встречается в пределах одной гаплогруппы, и обычно свидетельствует о серии бутылочных горлышек популяции, которая разошлась на ветви в древности, и претерпела трудное выживание в относительно недавнее время. О том, что исходные общие предки были в далекой древности, свидетельствуют масштабные различия между ветвями.
Если все гаплотипы дерева перемешать и усреднить, что будет совершенно неправильно, то фантомный
“общий предок” якобы жил примерно 4300 лет назад. Делать это неправильно потому, что дерево состоит из ветвей, каждая из которых описывает отдельную ДНК-генеалогическую линию, причем ветви - разного размера, то есть разного
“веса”. При усреднении теряются самые древние ветви, и дерево “омолаживается”. В “методе Животовского” это умоложение искусственно
“выправляется компенсацией” за счет умножения возраста почти в три раза. Почему именно на три? Да так. Так показала теория для 10 тысяч гаплотипов при их полной гомогенизации за всю историю человечества, что совершенно искусственное и нереальное допущение. Причем самое
“тупое” - это что умножение на три не зависит от формата гаплотипов, будь это 6- маркерные, 9-маркерные, 12-маркерные или 17-маркерные. Или любые другие. То есть принимается, что константа скорости мутации не зависит от того, какие маркеры в гаплотипе и столько их там. Уму непостижимо, что никто из
“популяционных генетиков” не обращает на это никакого внимания. Хоть бы кто подумал, что такого быть просто не может. Вот что такое отсутствие адекватной научной школы. Полная атрофия мышления. Короче, при использовании”метода Животовского” получится, что до общего предка всего дерева 12,800 лет. На самом деле этого нет и близко.
Рассмотрим дерево гаплотипов на рис. 1 с точки зрения ДНК-генеалогии.
Kangars/Turgeshes, modern Tubalars and Chelkans
Восемь гаплотипов на верхней левой ветви имеют базовый гаплотип (в формате Y-файлера, то есть DYS 393, 390, 19, 391, 385a, 385b, 439, 389-1, 392, 389-2 – 458, 437, 448, GATAH4, 456, 438, 635):
13 24 16 9
12 14 10 14 11 18
– 14 14 20 12 17
11 23 (No 57, 60-62, 68, 70, 71, 74)
1521
В ветви всего 10 мутаций (в отмеченных маркерах; в остальных маркерах мутаций вообще нет), что дает 10/8/0.034 = 37 → 38 условных поколений, то есть 950±315 лет до общего предка (стрелка – поправка на возвратные мутации, 0.034 – константа скорости мутации для 17-маркерных гаплотипов). Этот гаплотип заметно отличается от базового гаплотипа Русской равнины (на 9 мутаций)
13 25 16 11 11 14 10 13 11 17 – 15 14 20 11 16 11 23
и от базового гаплотипа юго-восточного субклада L342.2 (на 8 мутаций)
13 25 16 11 11 14 10 13 11 17 – 15 14 20 12 16 11 23
Это помещает их общего предка на 7250 лет назад (на 6750 лет назад с общим предком для субклада L342.2). Это не мог быть
“индоевропейский” R1a, в те времена они (the R1a “Indo-Europeans”) обитали в Европе, и даже на Русскую равнину еще не прибыли. Это – автохтонный общий предок, скорее всего фантомный (то есть датировка может быть заметно занижена), но это не
“индоевропейский” общий предок. Современную довольно молодую популяцию представляют в основном тубалары и челканы
(Turgeshes). Сейчас они монголоидны и тюркоязычны.
|
Tubalars and Chelkans are descendents of the Turgesh tribe, aka Kengeres and Kergej, supposedly a
descendent of the Kangar/Kangju state. In 651 -766 Turgeshes established their own Kaganate, they
suffered great losses in the wars of independence against encroaching China and pretentions of the Türkic
Kaganates, they carried a brunt of the Arab conquest campaigns in 711 - 724, suffered from
internecine wars of succession, and finally in 766 were dismembered by Karluks (Uigurs), who
displaced them from the Jeti-su area. During their independence, Turgeshes suffered numerous
genocides, and vanished as a great power. A part of the Kangars/Kangly retained their state, known
as Kangar Union (659-750) northwest of the Jeti-su, but eventually had to escape to the N.Pontic
(under the name of Bechens/Bajanaks/Pechenegs, 750-990), and then to Balkans, establishing Hrvatia/Croatia
and Bosnia-Hercegovina. A second recent bottleneck is connected with the 13th c. Mongol expansion,
when numerous Türkic tribes under Mongol flag campaigned and settled in the China, and E.Europe,
considerably reducing their presence in the Altrai, S.Siberia, and Middle Asia. Genetically, it is reflected in replacement in the E.Europe of the 2/3 eastern mtDNA with the western mtDNA
haplogroups.
The last bottleneck is connected with the Teleut Tele tribes escaping to the then-Russian Altai
fleeing form the army of the Manchurian-ruled Chinese Empire Qing exterminating Jungars and their
former kishtyms without any exceptions (events of 1750s - 1790s), bringing nearly complete extermination
of the Tele people, when no more than 10% of the Tele-confederated people survived
and found refuge in the mountainous areas in the Altai, Sayans, Pamir, and Chinese Turkestan.
Due to the pinpoint accuracy achieved by the DNA genealogy, we have a chance to peek into the
male and female genetic composition of the Kangars/Turgeshes, and trace their lineage in the modern
diaspora. The supposition that Kangars/Turgeshes populated Altai before or after displacement of
population in the Altai is unjustified, the Kangar phenotype and ontological descriptions, their
Jetyasar archeological culture, their kurgan burial customs, their popular cranial deformation
traditions, and even their coinage are detailed in the literature and can be traced across Eurasia.
The Altai findings reflect not only a different epoch, they also belong to the different
geographical area as much as the DNA of the modern Caucasoid Australians describes DNA of a different
geographical area.
|
Potentially Türks/Ashina Türks, or their parent tribe of Saka,
a part of modern Altai-kiji
У подножия описанной ветви находится мини-ветвь из четырех гаплотипов, три из которых одинаковы, и четвертый (56) отличается всего на одну мутацию (в DYS390).
13 26
16 10
11 14 10 14
11 18
– 15 14 21 12
16 11 23 (№ 27, 28, 29)
13 25 16 10 11 14 10 14 11 18 – 15 14 21 12 16 11 23 (№ 56)
|
Of these 4 branches, 3 may belong to the second-largest component of the Altai-kiji, the Türks/Ashina
Türks 22%, or their parent tribe of Saka/ |
Шесть мутаций (отмечены) разводит общих предков этих гаплотипов и базового гаплотипа Русской равнины на 6/0.034 = 176 → 214 поколений, или 5350 лет, и помещает ИХ общего предка на (5350+4600)/2 = 4975 лет назад. Это тоже слишком рано для прибытия на Алтай
“индоевропейских” R1a. Они (“Indo-Europeans”) прибыли на тысячу лет позже
(the“Indo-Europeans” in quotation marks are neither Indo- nor Europeans,
the label comes from the attribution by the latter days seers of the Andronovo Kurgan culture to the
Ossetians aka Iranians aka Indo-Europeans aka most reverend ourselves).
Одна мутация между приведенными выше гаплотипами помещает мини- ветвь на 1/0.34 = 29 → 30 поколений, то есть примерно на 750 лет назад. Но между этой мини-ветвью (алтай-кижи) и ветвью тубаларов и челканов
(Turgesh branch) – 6.5 мутаций, или 5900 лет между их общими предками. Это помещает ИХ общего предка на (5900+ 950 + 750)/2 = 3800 лет назад.
|
The indiscriminate blend of the Tele and Saka within the Altai-kiji blend, and Kangars/Turgeshes
genetically separated 3800 ybp, at about the time when the E.European agricultural refugees
just started their march southeast toward the Indian peninsula, and the Zhou Kurganians of South
Siberia reached the prehistoric China. |
Иначе говоря, от ДНК-линии, идущей от 3800 лет назад (которая в свою очередь тоже явно прошла бутылочное горлышко популяции) остались две подветви с возрастом 950 и 750 лет назад. Но расстояние между ними выдает, что они далеко разошлись от ИХ общего предка, и это расстояние можно без труда вычислить.
1522
Итак, мы видим, что верхняя левая часть дерева – это не “индоевропейские”, а автохтонные гаплотипы группы R1a. Они древние по происхождению, значительно древнее гаплотипов R1a (точнее, их общих предков) на Русской равнине.
Teleut Tele
Подобные расчеты позволяют получить следующие базовые гаплотипы и возраст общих предков ветвей. Малая ветвь слева из семи гаплотипов (в которых всего три мутации):
13 25 16 11 11 14 10 14 11 18 – 15 14 21 10 16 11 23 (325 лет до общего предка)
(No 3 - 7, 42, 46, 50).
|
325 years nearly agrees with the remnants of the Teleut Tele tribes escaping to the then-Russian
Altai fleeing form the army of the Manchurian-ruled Chinese Empire Qing exterminating Jungars and
their former kishtyms without any exceptions (events of 1750s - 1790s), bringing nearly complete extermination
of the Tele people, when no more than 10% of the Tele-confederated people survived
and found refuge in the mountainous areas in the Altai, Sayans, Pamir, and Chinese Turkestan. |
Kumans (Kumandins)
Довольно древняя ветвь из 6 гаплотипов (на 7 часов) [27 мутаций]:
13 25 15 10 11 14/15 10 13 11 17/18 – 15 14 20 12 16 11 23 (3800 лет до общего предка)
(No 8, 51, 59, 64, 65, 72)
Это вполне может быть субклад L342.2, базовый гаплотип которого на западе Русской равнины следующий:
13 25 16 11 11 14 10 13 11 17 – 15 14 20 12 16 11 23
|
This is an allusion to the “Indo-European” Andronovo presence, which is well-established as long as
it is within quotation marks, and to the linguistically “Indo-European” nature of the Andronovo
Kurganians, which is a conflicted concept with far-flung consequences. The ethnological assessment
strikingly conflicts with the linguistic assertions. Ethnologically, the Ural/Siberia migrants,
anthropologically defined as South Eurasian Anthropological Phenotype, and the “Indo-European” aka
“Aryan” southeast migrants were completely incompatible, with the main aspect being in their
relationship to the Kurgan Culture: the aka “Aryan” southeast migrants had none of it, as defined by
the compendium of the archeological traits, plus the aka “Aryans” had agricultural vs. Kurgan animal
husbandry economies, plus archeologically inferred organization of society, plus archeologically and
anthropologically established marital relationships, plus aka “Aryan” religious ideology vs.
etiology expressed in the kurgan tradition, plus markedly different phenotype constitution, plus
opposing lactose tolerance. It is not inconceivable that such disparate societies would speak the
same language, but the only asserted argument for that is the male Y-DNA, while not only the
mother's language is carried over by the female mt-DNA, but also the same Y-DNA studies assert that
Kurgan intrusions into Europe were linguistically non-Indo-European. Even unstoppable Mallory, who
sees Indo-European in every Caucasoid, found it extraordinary difficult to connect Andronovo with Aryan
migration to the northern India. He might find it even less acceptable that he himself
belongs to the “Erbin” stem, converted to the “Indo-Europeism” only lately in the 19th c. If further research would confirm the L342.2 subclade in the Altai
and in the Andronovo culture zone, it would necessitate a re-assessment of the L342.2 lineage
in respect to its presumed linguistic, biological, and ethnological connections. |
Probably, Tele
Молодая ветвь из 10 гаплотипов внизу дерева, в которой всего 4 мутации:
13 25 16 11 11 14 10 14 11 18 – 15 14 21 12 17 11 23 (300 лет до общего предка)
(No 25, 26, 34 - 41)
|
This could be the same migration of 1750s - 1790s, with a different founder |
Probably, Tele
Ветвь из семи гаплотипов в верхней правой части дерева, в которой всего 5 мутаций, что дает 5/7/0.034 = 21 поколение, или примерно 525 лет до общего предка:
13 26
16 10
11 17 11 14
11 18
– 15 14 19 11 15
11 23 (No 43-45, 47-49, 53)
Она на 11 мутаций на 17 маркерах отличается от базового гаплотипа субклада L342.2 (отмечено выше),
13 25 16 11 11 14 10 13 11 17 – 15 14 20 12 16 11 23
что разводит их общих предков на 11/0.034 = 324 → 472 поколения, или 11,800 лет, и помещает общего предка L342.2 и алтайской ветви на (11800 + 4900 + 525)/2 = 8600 лет назад. С базовым гаплотипом Русской равнины общий предок жил 7700 лет назад. Это – опять автохтонные общие предки гаплотипов. В этой (и следующей) ветви у них характерный признак, DYS385 = 11-16, в то время как на Русской равнине он 11-14.
1523
Probably, Tele
Подветвь из 9 гаплотипов на 3 часа (в которой всего 7 мутаций, то есть общий предок жил 7/9/0.034 = 23 поколения, то есть 575 лет назад), с базовым гаплотипом
13 26 16 11
11 17 11 14 11 17
– 15 14 19 11 15 11 23 (No 1, 2, 9-11, 12, 15, 52, 54)
Видно, что это родственная ветвь предыдущей. У них одинаковые значения DYD385=11-17, и они отличаются всего двумя мутациями, то есть их общие предки расходятся на 2/0.034 = 59 → 63 поколения, то есть на 1575 лет. ИХ общий предок жил (1575+525+575)/2 = 1340 лет назад. Ясно, что эта двойная ветвь молодая (в отношении своего общего предка). Видно, как фрагментированы ветви, как они рассыпаются на совсем недавно выжившие и давшие недавних же потомков.
|
The timing and scatter resembles the problems at the end of the Western Türkic Kaganate (ca 660s),
whose Kagan Chjenchu was killed, and whose elite had to flee and hide, naturally in the Altai, from
the mobile nomadic armies assembled by the Tang ruler. The modern carriers would tentatively be the
descendents of the Türks/Ashina Türks, a second-largest component of the indigenous Altai-kiji on
the last of their numerous retreats to the Altai. But among numerous Türkic tribes of the time, not
only the Ashina Türks faced problems of survival, and each one of them is a potential candidate for
the fossil DNA. |
Пара гаплотипов (13, 14) одинаковы, поэтому технически их общий предок не имеет возраста:
13 25 16 11 11 18 11 14 11 17 – 15 14 19 10 15 11 23 (0 лет) (No 13, 14)
Kangars/Turgeshes, modern Tubalars and Chelkans
Наконец, последняя ветвь (на пять часов) из 6 гаплотипов, в которых всего 11 мутаций, то есть 11/6/0.034 = 54 → 57 поколений, то есть примерно 425 лет до общего предка, имеет базовый гаплотип
13 24/25 16 11 11 14 11 14 11 17 – 16 14 20 14 16 11 23
Он резко отличается от соседней двойной ветви выше по дереву, особенно по GATAH4 (отмечено). В целом отличия от базового гаплотипа Русской равнины составляют 6.5 мутаций
13 25 16 11 11 14 10 13 11 17 – 15 14 20 11 16 11 23
и от ветви выше по дереву – на 12.5 мутаций
13 26 16 10 11 17 11 14 11 18 – 15 14 19 11 15 11 23
что дает 12.5/0.034 = 368 → 570 поколений, или 14,250 лет между общими предками. ИХ общий предок жил (14250+ 525+425)/2 = 7600 лет назад. Это опять же автохтонный общий предок.
Если мы в итоге выпишем базовые гаплотипы всех десяти ветвей дерева, включая (кажущийся) базовый гаплотип всего дерева на вершине (последний гаплотип в матрице), получим следующую матрицу:
1524
13 24 16 9 12
14 10 14 11 18 – 14 14 20 12 17 11 23
13 25.5 16 10
11 14 10 14 11 18 – 15 14 21 12 16 11 23
13 25 16 11 11 14 10 14 11 18 – 15 14 21 10 16 11 23
13 25 15 10 11 14.5
10 13 11 17.5 – 15 14 20 12 16 11 23
13 25 16 11 11 14 10 14 11 18 – 15 14 21 12 17
11 23
13 26 16 10
11 17 11 14 11 18 – 15 14 19 11 15 11 23
13 26 16 11 11 17 11
14 11 17 – 15 14 19 11 15 11 23
13 24.5 16 11 11 14 11
14 11 17 – 16 14 20 14 16 11 23
13 25 16 11 11 18 11 14 11 17
– 15 14 19 10 15 11 23
13 25 16 11 11 14 10 14 11 18 – 15 14 21 12 16 11 23 |
(950 лет)
(750 лет)
(325 лет)
(3800 лет)
(300 лет)
(525 лет)
(575 лет)
(425 лет)
(0 лет)
(0 лет) |
Kangars/Turgeshes, modern Tubalars and Chelkans
Potentially Türks/Ashina Türks, or their parent tribe of Saka
Tentatively Tele
Kumans (Kumandins)
Tentatively Tele
Tentatively Tele
Tentatively Tele
Tentatively Tele
Tentatively Tele
Potentially Türks/Ashina Türks, or their parent tribe of Saka |
Вся матрица содержит 51 мутацию (отмечены), что помещает возраст общего предка на 51/10/0.034 =
150 → 176 поколений, или на 4400 лет ниже среднего возраста всех базовых гаплотипов матрицы (765
лет), то есть общий предок всех ветвей жил 4400+765 = 5165 лет назад. Это может соответствовать
возрасту афанасьевской культуры (Southern Siberia Nomadic pastoralism,
Southern Eurasian Anthropological Phenotype, 5700/5300 - 4500/4000 ybp, 3700/3300 - 2500/2000 BC).
Литература
Dulik, M.C., Zhadanov, S. I., Osipova, L.P., Askapuli, A., Gau, L., Gokcumen, O., Rubinstein, S., Schurr, T.G. (2012) Mitochondrial DNA and Y chromosome variation provides evidence for a recent common ancestry between Native Americans and indigenous Altaians.
Amer. J. Human Genetics, 90, 1-18.
DOI 10.1016/ajhg.2011.12.014 Gonzalez-Ruiz, M., Santos, C., Jordana, X., Simon, M., Lalueza-Fox, C., Gigli, E., Aluja, M., Malgosa, A. (2012) Tracing the origin of the East-West population admixture in the Altai region (Central Asia).
PLOS One, 7, 1-11. e48904
Klyosov, A.A., Rozhanskii, I.L. (2012) Haplogroup R1a as the Proto Indo- Europeans and the legendary Aryans as witnessed by the DNA of their current descendants.
Adv. Anthropol. 2, No. 1, 1-13.
Klyosov, A.A. (2009) DNA Genealogy, mutation rates, and some historical evidences written in Y-chromosome. I. Basic principles and the method. J. Genetic Genealogy, 5, 186-216.
Klyosov, A.A. (2009) DNA Genealogy, mutation rates, and some historical evidences written in Y-chromosome. II. Walking the map. J. Genetic Genealogy, 5, 217-256.
Klyosov, A.A. (2009) A comment on the paper: Extended Y chromosome haplotypes resolve multiple and unique lineages of the Jewish priesthood.
Human Genet. 126, 719-724.
1525


 , Булгарами (без Тохаров),
Хорезмийскими Оттокарами (Асы-Тохары),
Кыпчакскими Токс-оба (Токсобичи) в Северопонтике, Дюгерами и Дигорами по обе стороны Каспия, и
еще другими осколками. На востоке, Асы-Тохары известны как Аштаки (Кит. Ашиде, тамга
, Булгарами (без Тохаров),
Хорезмийскими Оттокарами (Асы-Тохары),
Кыпчакскими Токс-оба (Токсобичи) в Северопонтике, Дюгерами и Дигорами по обе стороны Каспия, и
еще другими осколками. На востоке, Асы-Тохары известны как Аштаки (Кит. Ашиде, тамга