|
Синопсис
Данная статья направлена на реконструкцию истории древних миграций маркера R1b между 16.000 и
1.500 лет назад (лн). Четыре тысячи четыреста восемь (4408) гаплотипов гаплогруппы R1b (с
субкладами) были рассмотрены с точки зрения базовых (наследственных) гаплотипов у населения
гаплогруппы R1b и расчетного времени до их общих предков. Рассматриваются области от Южной
Сибири/Центральной Азии на востоке (где гаплогруппа R1b возникла ~16000 лн) через Северный
Казахстан, Южный Урал до Восточно-Европейской равнины и далее на запад в Европу (северный маршрут
вступающих в Европу около 4500 лн); от Восточно-Европейской равнины на юг к Кавказу (6000 лн), Малой
Азии (6000 лн) и Ближнему Востоку (6000 - 5500 лн) к Балканам в Европе (южный маршрут вступающих в
Европу около 4500 лн); по Северной Африке и Средиземному морю (5500 - 5000 лн) через Египет до
Атлантического океана, на север до Иберии (Североафриканский маршрут с приходом на Пиренеи 4800 лн).
Повидимому, Арбины (носители гаплогруппы R1b) по пути маршрута их миграции на Ближний Восток и в
Южную Месопотамию создали Шумерскую культуру (и государство), двигаясь на запад в Европу (5000 -
4500 лн) в основном с субкладом R-M269 и его нижестоящим субкладом L23. Этот последний субклад почти
отсутствовал по Североафриканскому маршруту, и/или не выжил перехода к Иберию или не был
засвидетельствован позже. По прибытии в Иберию (4800 лн) от субклада M269 откололся субклад M51, и
вскоре после этого откололся нижестоящий субклад L11. Эти группы, ставшие известными как
Колоколовидные Кубки, двинулись на север, вместе с вновь возникшими субкладами P312 и L21 (которые
откололись в несколько следующих веков после P312). Эти субклады и их нижестоящие клады по сути, без
значительных перерывов, населили Европу (гладкие древья гаплотипов демонстрируют почти
безостановочное распространения гаплотипов R1b в Европе). Они зарегистрированы от Атлантического
океана на восток до Балканов, Карпат, сегодняшней Польши до западной границы Восточно-Европейской
равнины и до Балтийского моря. На островах история миграции R1b была другая. Носители L11, L21 и
P312 добрались на острова по суше и по морю одновременно с теми Арбинами, которые заполненяли Европу
между 4000 и 2500 лн и сформировали соответствующие “местные” субклады P314, M222 и L226, которые во
многом заселили острова. В результате значительная часть островов населена почти исключительно
Арбинами, частота которых достигает 85% - 95% среди нынешнего населения. В целом, частота Арбинов в
Западной и Центральной Европе достигает - хотя и не равномерно - около 60% населения. Это
исследование по существу представляет собой пример применения ДНК-генеалогии в деле изучении истории
человечества. Введение Из-за отсутствия общего названия для носителей
гаплогруппы R1b (с субкладами) и их языков в древние времена, которых несли тысячелетиями и в конце
концов принесли в Европу как не-Индо-Европейские языки, я их называю Арбины (от R1b), как людей, так
и их родные языки, по аналогии с Арийцами (от Аранов, по тому же формату как и Арбины), которые по
сути принадлежали к гаплогруппе R1a (Клесов и Рожанский, 2012а). Происхождение и история
гаплогруппы R1b, которых я называю Арбины, в настоящее время составляют почти 60% жителей Западной и
Центральной Европы. Кроме того, Арбины населяют значительную часть Кавказа, Анатолии и Малой Азии,
Ближнего Востока, и многие места в Центральной Азии, включая Южную Сибирь, Алтай, Туву,
Северо-Западный Китай, Среднюю Азию, некоторые области Урала и Среднего Поволжья с этническими
группами и населениям как Башкиры, Татары, Чуваши, и другие. История R1b была значительно искажена
с самого начала “генетической генеалогии” с конца 1990-х годов, когда было заявлено, совершенно
безосновательльно, что гаплогруппа R1b возникла в Европе около 30.000 лет назад (лн). Необоснованно,
потому что заявки действительно были основаны на отсутствии данных. Такие данные никогда не
существовали. Тем не менее, заявления и заявки, как “Около 30.000 лет назад, у потомока клана,
прокладывавшего себе путь в Европу, появился маркеру M343, определяющий маркер гаплогруппы R1b. Эти
путешественники являются прямыми потомками людей, которые доминировали в экспансии людей в Европу, а
именно кроманьонцев” (Spencer Wells, “Глубокое наследие”, 2006). Эта и подобные заявки, такие
как R1b (и его субклады M269) были “хорошо распространены по всему палеолиту Европы”, “одновременно
с Ориньякской культурой”, “самая ранняя экспансия в Европу в верхнем палеолите ~30.000 лет
назад” Уэллса (Wells), Семино (Semino), Андерхилл (Underhill), Кавалли-Сфорца (Cavalli-Sforza),
Чиниоглу (Cinnioglu), Кивислид (Kivisild), Виик (Wiik), и многих других (напр. Semino et al., 2000;
Wells et al., 2001; Cinnioglu et al., 2004; Wiik, 2008) по сути опирались на “мысль”, что если люди
жили в Европе около 30.000 лет назад, они обязательно принадлежали к гаплогруппам R1b, а не I, G, J,
E, F или любым другим гаплогруппам. Были ли проанализированы хоть какие-то гаплотипы? Их мутации
были просчитаны? Сделаны хоть какие-то хронологические оценки? Ничего из вышеперечисленного не
существовало.
87 "Палеолитическое происхождение” R1b в Европе, или их
“Палеолитические миграции” в Европу около 10000 - 8000 лн по-прежнему утверждаются в последних
научных работах, таких как (Myres et al., 2010; Balaresque et al., 2010; Morelli et al., 2010). Эти
посылки и в последуюие расчеты, как правило, основаны на "скорости мутаций населения”
(Животовский/Zhivotovsky et al., 2004;. Hammer et al., 2009; Underhill et al., 2009), которые обычно
преувеличивают хронологическую оценку миграций и событий на 200% - 400%, так как они основаны на
грубом, искусственном и нереально наивном и обобщенном обосновании (см. критику Клесов, 2009a,
2009b, 2009c; Рожанский и Клесов, 2011). “Научные работы”, размещая происхождение R1b в Малую Азию
или поблизости, не рассматривали области к востоку от Малой Азии, а также гаплотипы R1b этих
восточных областях. Короче говоря, вся картина миграций R1b и их история в находятся в беспорядке, и
"популяционные генетики” продолжают продвигать неверные выводы из-за их методологии. Методология
данного исследования/анализа включает в себя рассмотрение Y-хромосомы удлиненных 67 и 111-маркерных
гаплотипов, если таковые имеются. Методология была подробно описана в предыдущих работах в этом
журнале (Рожанский и Клесов, 2011; Клесов и Рожанский, 2012A) и в других местах (напр. Клесов,
2009а, 2009с, 2009d), и в Материалах и Методах этой статьи. Как описано (Клесов и Рожанский,
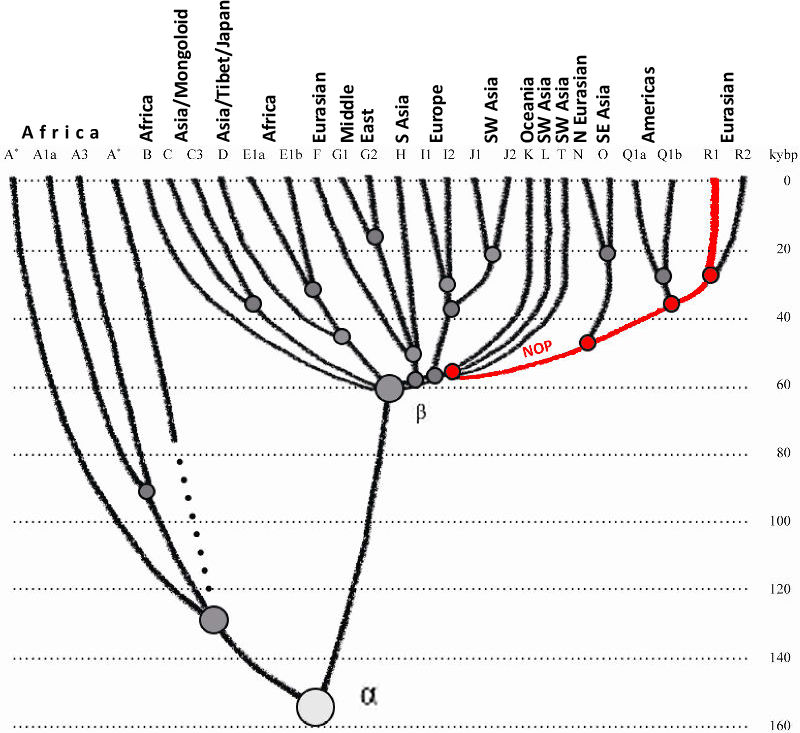
2012b), Европеоиды (Кавказоиды) появилась ~58.000 лн. Они постепенно разветвились по нижестоящим
гаплогруппам и их субкладам, и мигрировали на север, запад, юг и восток. Гаплогруппа NOP, которая
была среди них, возникли ~48.000 лн, и двинулись на восток, предположительно в сторону Южной Сибири
и/или прилегающих областей. Гаплогруппа P отделилась ~ 38.000 лн, предположительно, в Южной Сибири,
и произвела гаплогруппы R и потом R1 ~30.000 - 26.000 лн (см. схему в Клесов и Рожанский, 2012b).
Гаплогруппа R1b возникла ~16000 лн, как это будет показано далее в этой статье.
Рис 3 из Клесов, А. и Рожанский, И. (2012). Пересматривая Теорию “Из Африки” и происхождение
Европеоидов (Кавказоидов) в свете ДНК-генеалогии
Advances in Anthropology, (Достижения в антропологии), 2, 80-86 |
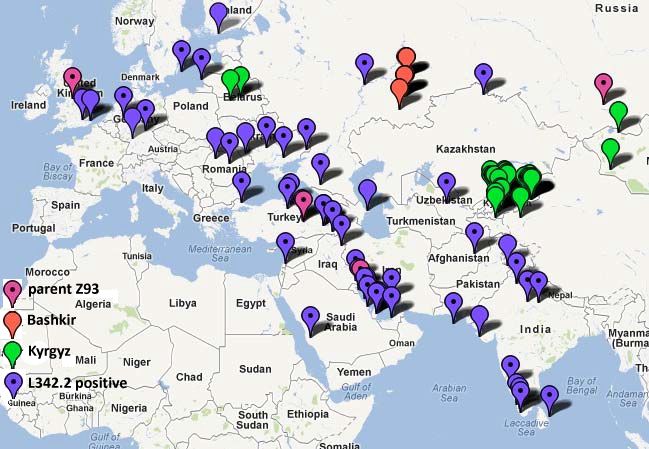
Хронология может быть восстановлена из серии гаплотипов R1b, доступной из баз данных (см.
Приложение). Самый дальние гаплотипы R1b (проявляющие наибольшие мутационные отличия)
Европейских гаплотипов R1b были найдены в населениях Сибири и Средней Азии. Среднеазиатские носители
гаплотипа R1b имеют самых древних общих предков с европейскими носителями R1b, и те древние общие
предки жили ~16000 лн в Центральной Азии. Мы не знаем пока то ли в Южной Сибири или Средней Азии;
однако доказательства показывают, что это было где-то в той огромной области.
Следующий синипсис по истории гаплогруппы R1b взят из Клесов А. (2011), Гаплотипы R1b1a2-P312 и
относящиеся субклады: происхождение и “возраст" самых недавних общих предков//
Вестник
Русской Академии ДНК-генеалогии , Vol. 4, № 6, 2011. Синипсис о гаплогруппах, сроках,
миграции, и языке:
1130ИСТОРИЯ ГАПЛОГРУППЫ R1b
от Центральной Азии до Европы
Появление R1b в Центральной Азии (Алтай, В.Туркестан, Северо-Западный Китай), ее древние миграции в
Европу; Башкиры, народы Кавказа, Анатолия, Шумер, Северная Африка, Иберия, Баски,
континентальная Европа, Британские острова), Колоколовидные Кубки как R1b1a2-P312 и U106- и их
вышестоящие (L51, L11/L151/P310/P311) и нижестоящие субклады в Европе, и не-Колоколовидные Кубки
R1b1a2-M269, -L23, и их нижестоящим субклады в Европе
Как было показано в серии более чем 30-ти работ (Клесов, 2008-2011, см. выше и список литературы
в конце статьи), гаплогруппа R1b1, а также ее сестринская гаплогруппа R1a1 возникли в Центральной
Азии (Южной Сибири и сопредельных областях, таких как Алтай, В.Туркестан включая Северо-Западный
Китай). R1a возникла около 21.000 лет назад (лн), потом возникла R1b1, около 16.000 лн. Так как
самый медленный (по отношению к мутациям) 22-маркерный европейский базовый гаплотип R1a1-M17
(предположительно предковый) [DYS 426, 388, 392, 455, 454, 438, 531, 578, 395S1a, 395S1b, 590, 641,
472 , 425, 594, 436, 490, 450, 617, 568, 640, 492]
12 12 11 – 11 11 – 11 – 11 8 17 17 8 10 8 12 10 12 12 8 12 11 11 12 (R1a)
и базовый гаплотип 22-маркерный R1b1a2-M269
12 12 13
– 11 11 – 12
– 11 9 15 16
8 10 8 12 10 12 12 8 12 11 11 12 (R1b1a2)
отличаются на 7 мутаций, как отмечено выше (Клесов, 2011a), что дает 7/0.006 = 1167 поколений
без поправки на обратные мутации, или 1380 поколений с коррекцией (поправочный коэффициент в данном
случае равен 1,187, см. стр. 1280-1281 об этом вопросе), то есть 34.600 лет между их общими
предками, ИХ общий предок жил примерно (34600 +5000 +7000)/2 = 23.300 лет назад. Это общий предок
гаплогруппы R1. Здесь 0,0060 является константа скорости мутации для 22-маркерного гаплотипа
(мутация/гаплотип/поколение 25-ти лет) [Клесов, 2011b, и статья по этому вопросу, стр. 1239-1256].
Действительно, если общий предок R1 жил 23.300 лн, то базовый гаплотип R1a1-M17 в Восточной
Европе появился (2.300 + 16.000) = 18.300 лет после появления R1, и R1b1b2-M269 возник (7.300 +
9.000) = 16.300 лет после появления R1; следовательно, разница между двумя базовыми гаплотипами выше
равна (18.300 + 16.300) = 34.600 лет, точно как показывают два 22-маркерных базовых гаплотипа с
разницей в семь мутаций между ними.
После того как гаплогруппа R1 возникла около 23.000 лет назад, и ее нижестоящая R1b возникла в
Центральной Азии 16.000 лет назад, R1b мигрировала через Северный Казахстан (включая позжую
Ботайскую археологическую культуру, 6500-5500 лн), через территорию современных Башкир (13-11-8
тысяч лн), создав Сероглазово археологическую культуру (13-11 тысяч лн), а затем Средневолжские
археологические культуры (Средневолжскую культуру 8-7 тысяч лн; Самарскую культуру 7.000 лн;
Среднестоговую культуру 7.500-5.500 лн; Хвалынскую культуру 7-6 тысяч лн; Камскую культуру 7-5 тысяч
лн), то есть культуры (ориентировочно) названые “Pre-Курганные культуры”; субклад R1b1a2-M269
возник около 7000 лн, а затем возник субклад R1b1a2-L23, около 6500 лн. Они обе пришли в область
Кавказа около 6500-6000 лн, и возможно раннее. Те R1b1b2 говорили на не-Индо-Европейском языке
(языках), которые могут быть смутно сейчас прослеживаеться под разными названиями. Некоторые
называют их Прото-Тюркским языком, некоторые Сино-Кавказским языком, некоторые Палео-Евразийскм
языкм, некоторые называют их язык Ербинским (из-за гаплогруппы R1b их носителей), некоторые называют
их Западными Кавказскими или Северо-Западными Кавказскими языками, некоторые называют их Баскским
языком, и находят некоторое сходство между Кавказскими и Баскским языками, и т.д.
1131
На самом деле, это сходство действительно существует, хотя и в довольно слабой форме, потому что
с тех пор прошли тысячи лет. Тем не менее, в настоящее время большинство гаплотипов R1b1a2 на
Кавказе (в Армении, Дагестане, Грузии) относятся к древнему субкладу L23 (с общим предком на Кавказе
около 6000 лн), и имеют характерный аллеле DYS393 = 12, в отличие от DYS393 = 13 в большинстве
европейских гаплотипов R1b1b2.
С Кавказа, носители R1b1a2-L23 и R1b1a2-M269 отправились через горы на юг, в Анатолию (общий
предок 6000 лн), а затем разделились на три основные
маршрута. Один пошел далее на юг, к рекам Тигру и Евфрату, и они стали Шумерами. Многие современные
Ассирийцы, потомки древних Шумеров, до сих пор имеют их гаплотип R1b1a2. Другой пошел на запад,
через Малую Азию, и пришли в Европу, на Балканы и область Средиземноморья около 4500 лн. Третья
группа пошла через всю Северную Африку и Египет (и, кстати, возможно оставила там некоторых R1b1b2
фараонов) к Атлантике и пошел через Гибралтар на Пиренейский полуостров около 4800 лн. Они стали
Колоколовидными Кубками, и продвинулись на север в континентальную Европу. Культура Колоколовидных
Кубков в Европе длилась примерно от 4400 до 3800 лн.
По-видимому, носители R1b1a2 пришли на Пиренеи главным образом как субклады L51/M412 и/или
L11/L151/P310, и вскоре разделились там на нижестоящие субклады U106 и P312, которые отправились в
континентальную Европу как Колоколовидные Кубки выше, с нижестоящими субкладами как U198, U152, L2,
L20, L4, L21, M222, L226, SRY2627, и т.д. Позади они оставили Басков, которые все еще сохраняют свой
древний Ербинский (R1b) язык, похожий по типу с некоторыми Кавказскими языками, и с Шумерским
языком, и имеют некоторыми элементы Прото-Тюркских языков Азии и далее Сибири.
До начала 1-го тысячелетия до нашей эры язык R1b1a2 в Европе был преимущественно (или только)
не-Индо-Европейским (не-ИЕ). Не существует ни одного твердого
доказательства иного, что R1b1a2 в Европе говорили на ИЕ языках до 1-го тысячелетия до нашей эры.
Например, Кельтские языки, а также “Прото-Кельтские” языки, помещаются лингвистами не ранее чем
через 800-900 гг. до н.э., и как правило, все более ранние опреселения беспочвенны. В то же
время мы знаем, что это носители R1a1 принесли свои ИЕ языки в Индию около 3500 лн, и R1a1
датирована в Европе по крайней мере 6000 лн. В Европе не существовало никаких R1b1a2 в те времена. В
Европе 6000 лн и пожалуй ранее, говорящими на прото-ИЕ языке (языках) должны были быть носители
R1a1.
1132
Корни Колоколовидных Кубков можно проследить, сравнив 67-маркерный базовый гаплотип современных
потомков “параллельных” субкладов U106 и P312:
13 23
14 11 11 14 12 12 12 13 13 29 – 17 9 10 11 11 25 15 19 29 15 15 17 17 – 11 11 19 23
16
15 17
17 37 39 12 12 – 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12
12 15 8 12 22 20 13 12 11 13 11 11
13 12 (U106)
13 24 14 11 11 14 12 12 12 13 13 29 – 17 9 10 11 11 25 15 19 29 15 15 17 17 – 11 11 19 23 15 15
18 17 36 38 12 12 – 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11
12 12 (P312)
Между ними есть шесть мутационных отличий (отмеченых выше в базовм гаплотипе U106), что
соответствует 6/0.12 = 50 → 53 “условных” поколений (25 лет на поколение), то есть 1325-летний
промежуток времени между их общими предками. Знак → означает поправку на обратные мутации (см.
ниже). Поскольку общий предок U106 и P312 жил 4175±430 лн и 4100±415 лн соответственно (см. ниже, а
также 3950±400 лн, Клесов, 2010а, что практически та же цифра в пределах погрешности), ИХ общий
предок жил (1325+4175+4100)/2 = 4800 лет назад. Это достаточно хорошо согласуется со временем
Колоколовидных Кубков.
ГЕНЕТИЧЕСКOЕ РЕЗЮМЕ
Подробный анализ 2299 67-маркерных гаплотипов набора, предоставленного Майкл Уолш (Michael
Walsh), администратором сайта R1b1a2-L21
http://www.familytreedna.com/public/R-L21/default.aspx?section=yresults позволил
идентифицировать базовые (наследственные) 67-маркерные гаплотипы и время до их общих предков
следующим образом
(схема недостаточна в отношении Азиатских субкладов):
| • | • | • | • | • | R1b1a2 |
| | | | L265, M269, M520, S3, S10,
S13, S17 |
~7000 лн |
Азия |
| • | • | • | • | • | • | R1b1a2a |
|
|
|
|
| L23/S141, L49.1 |
~6200 лн |
Азия |
| • | • | • | • | • | • | • | R1b1a2a1a |
|
|
|
| L51/M412/S167 |
5300±700 лн |
Азия |
| • | • | • | • | • | • | | • |
R1b1a2a1a1 |
|
| | |
L11, P310, P311 |
~4800 лн
(алт. 4575±580 лн) |
Европа |
| • | • | • | • | • | • | | • | • |
R1b1a2a1a1a | |
|
|
M405/S21/U106 |
4175±430 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
null mutation U106 | |
|
|
3325±450 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
R1b1a2a1a1b |
|
|
P312/S116 |
~4800 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
null mutation P312 | |
|
3575±400 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b1 M65 | |
|
~1800 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
R1b1a2a1a1b2 |
| |
M153 |
~3640 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
R1b1a2a1a1b3 |
| |
S28/U152 |
4125±450 лн
(алт. 3800±380) |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
null mutation U152 |
|
| 3525±460 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b3c |
|
L2/S139 |
4025±410 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
| |
R1b1a2a1a1b3c1• | L20 |
3650±400 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b3d |
|
L4/S178 |
1275±290 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
R1b1a2a1a1b4 |
| |
L21/M529/S145 |
3750±380 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
null mutation L21 |
|
| 3025±460 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
| |
null mutation L21 | |
1500±325 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
| Unidentified clade | |
|
| 1650±175 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b4b |
|
M222 |
1450±160 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b4e |
|
L144 |
~4000 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b4f |
|
L159.2 |
1775±200 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b4g |
|
L193 |
1275±170 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b4h |
|
L226 |
1500±170 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b4i |
|
P314.2 |
2225±300 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
R1b1a2a1a1b5 |
| |
L176.2/S179.2 |
3675±560 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b5a |
|
SRY2627 |
3150±320 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
| |
null mutation SRY | |
~2800 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
|
R1b1a2a1a1b5b |
|
L165/S68 |
~3000 лн |
Европа |
| • | • | • | • | • | • | | • | • | • |
R1b1a2a1a1b6 |
| |
L238/S182 |
~1000 лн |
Европа |
|
Здесь термины “гаплогруппа” и “субклад” используются как почти эквивалентные, потому что все
гаплогруппы по существу являются субкладами их вышестоящих гаплогрупп, и использование того или
другого термина идет по контексту. Это показано на следующей диаграмме (ISOGG-2012, фрагмент
http://www.isogg.org/tree/ISOGG_HapgrpR.html), который относится к наиболее древним субкладам
гаплогруппы R1b:
| R1b | | | | M343 |
| | R1b1 | | | L278, M415, P25_1, P25_2, P25_3 |
| | | R1b1* | | - |
| | | R1b1a | | P297, L320 |
| | | | R1b1a1 | M73, M478 |
| | | | R1b1a2 | L265, M269, M520, S3, S10 |
| | | R1b1b | | M335 |
| | | R1b1c | | V88 |
| | | | R1b1c* | - |
| | | | R1b1c1 | M18 |
| | | | R1b1c4 | V69 |
Взгляд с высоты птичьего полета на гаплотипы R1b, и самые древние популяции R1b
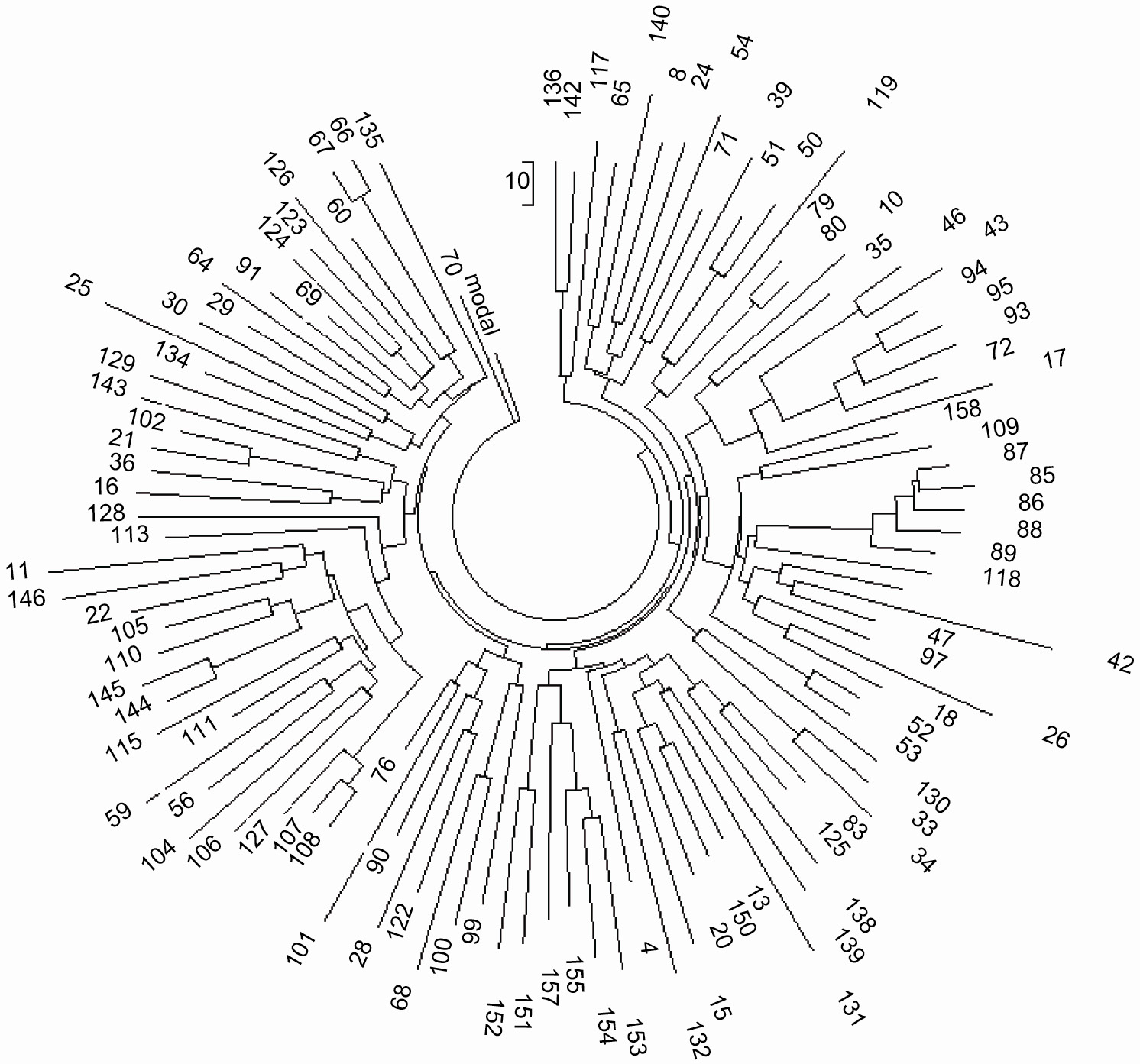
Рисунок 1 дает обзор древа гаплотипов R1b с 338-ю гаплотипами в кратком 8-маркерном формате. Цель
показа - показать сложную картину гаплогруппы R1b и определить наиболее древнюю ветвь на древе.
Древо гаплотипов показаное ниже составлено с помощью компьютерной программы которая обьединяет
ветви на основе сходства их аллелей в соответствующих маркерах (или локусах) в Y-хромосоме, и
динамики их аллелей (Клесов, 2009с и ссылки в ней). Гаплотипы, идентичные друг другу и преобладающие
в наборе данных, находятся на вершине древа; в относительно недавних наборах данных они обычно
означают “базовые” или “предковые” гаплотипы, из которых все остальные гаплотипы выводятся через
мутации. Ветви расположеные близко к “стволу” древа содержат немного мутаций от базового гаплотипа,
следовательно, они являются относительно “молодыми". Древнейшими ветвями являются те, которые
отходят от ствола, так как в их гаплотипах они содержат большинство мутаций.
Рисунок 1. Древо 338-и 8-маркерных гаплотипов R1b1, состоящие из данных, приведенных в (Zhong
et al., 2010).
Дерево содержит гаплотипы субклада R1b* (8 гаплотипов), R1b1* (2 гаплотипа), R1b1c (2 гаплотипа),
R1b (140 гаплотипов), R1b1a1 (12 гаплотипов), R1b2b (21 гаплотип), R1b1a2 (153 гаплотипов). В
отношении этнического происхождения и/или места, гаплотипы принадлежат 40 западноевропейцам, 17
Иберам, 84 Туркам, 48 Албанцам, Македонцам и Румынам, 40 Итальянцам, 30 Критянам, 13 грекам и
Македонцам, 9 Румынам из другого региона, 13 Пакистанцев, 4 Индусам, 2 Японцам, 2 Монголам
(один из них, № 176, относится к субкладу R1b1b1-M73 и выделяется в нижней части древа), 2 Тибетцам,
1 Кыргызам из Синьцзяна, 7 Китайцам, 16 Уйгурам (12 из В.Туркестана/провинции Синьцзян, шесть из них
относятся к субкладу R1b* , три R1b1b1 и три R1b1b2), четыре Китайцам (три R1b2b и один R1b1b2), 2
Китайцам племени Хуэй (Hui) (в основном Uйгуры, так как Kитайцы используют
этноним Хуэй = Уйгур для общего обозначения Мусульман), и 6 Китайцам других
национальностей (Hazak R1b1b1, Ту R1b2b , и четыре Наси (Naxi) R1b2b) |

На Рисунке 1 сравнительно недавняя и преобладающе R1b ветвь имеет базовый гаплотип
13 24 14 11 Х Х Х 12 Х 13 13 29.
Тридцать пять из их идентичных гаплотипов находятся на вершине древа. Здесь X замещает
отсутствующие аллели в гаплотипе представленом в стандартном двенадцати-маркерном формате FTDNA
(определения см. Материалы и Методы). Этот базовый гаплотип принадлежит к числу самых
густонаселенных Европейских базовыми гаплотипами субклада R-M269, который в свою очередь включает в
себя субклад R-P312 и многие его нижестоящие субклады. Например, базовый 67-маркерный субклад
гаплотипа P312 выглядит следующим образом (Клесов, 2011b):
13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 -
11 11 19 23 15 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20
13 12 11 13 11 11 12 12 (P312)
Аллели, идентичные очевидно “базовому” гаплотипу в древе на Рисунке 1, выделены жирным.
Действительно, субклад R1b1a2-M269 занимает большую часть древа на Рисунке 1, так как "R1b”,
как указано в перечне, в основном также R-M269.
Анализ древа показывает, что многие Азиатские гаплотипы R1b, в частности, все Индийские,
большинство Пакистанских, четверть Уйгурских, один Японский, один Тибетский, и два Китайских Хань не
являются “коренными” гаплотипами, но такими, которые тесно связаны с Европейскими гаплотипами и
делят с ними тем же ветви (Клесов, 2010а). Другими словами, они либо являются “возвращенцами” в свои
регионы вместе с Европейцами, или имеют происхождение Европейского R1b. Они неразличимы от
Европейских гаплотипов R1b.
Некоторые из гаплотипов, однако, образуют отдельные ветви. Самой замечательной ветвью является
наиболее удаленная от ствола, а значит самая древняя, она расположена в левой верхней части древа.
Она содержит 12 гаплотипов, которые принадлежат к субкладам R1b1a1-M73 и R1b-M343, взятым у Уйгуров
и близких им племен Наси (Naxi), Хань и Ту
(Zhong et al., 2010). Все 12 гаплотипов являются производными от базового гаплотипа
13 24 15 11/10 XXX 12 X 14/13 12 30,
и коллективно содержат 65 мутаций от него. Используя “линейную” формулу, мы получаем 65/12/.013 =
417 → 619 условных поколений (стрелка указывает коррекцию на обратные мутации), или 15.475±2500 лет
от общего предка (расчет поясняется в разделе Материалы и Методы). Эта дата находится в хорошем
согласии с 16.000±1400 лет от общего предка гаплотипов R1b (Клесов, 2008a, 2009d) и будет
дополнительно подтверждена позднее в этой статье. Это также указывает, что гаплогруппы R1b возникла
в Центральной Азии, и видимо в области Алтая в Южной Сибири, где их вышестоящие гаплогруппы NOP → NO
+ P; P → R + Q; R → R1 + R2; R1 → R1a + R1b все мигриривали и там жили в течение периода времени
между 52-м и 20-15 тысячелетий назад (Клесов и Рожанский, 2012b).
Сибирские, Башкирские, и Центральноазиатские гаплотипы R1b-M73
Хотя субклад M73 расположен в нескольких шагах от верха диаграммы, он содержит самые отдаленные
известные гаплотипы Европейцев, как географически, так и по их мутациям. Рисунки 2 и 3
показывают два древа гаплотипа M73 из двух различных наборов данных, один был получен
непосредственно от исследователей и опубликован ранее (Клесов, 2008а), второй был собран из проекта
гаплогруппы R1b1b1 (см. Рисунок 3 и ссылки). Оба они имеют принципиально одинаковую форму, с
двумя основными ветвями на Рисунке 2 и тремя на Рисунке 3.
| Рисунок 2. Древо 14 25-маркерных гаплотипов для гаплотипа R1b1-M73 Центральной Азии (Узбеки,
Таджики, Тувинцы, Уйгуры, Казахи) (Клесов, 2008а) |
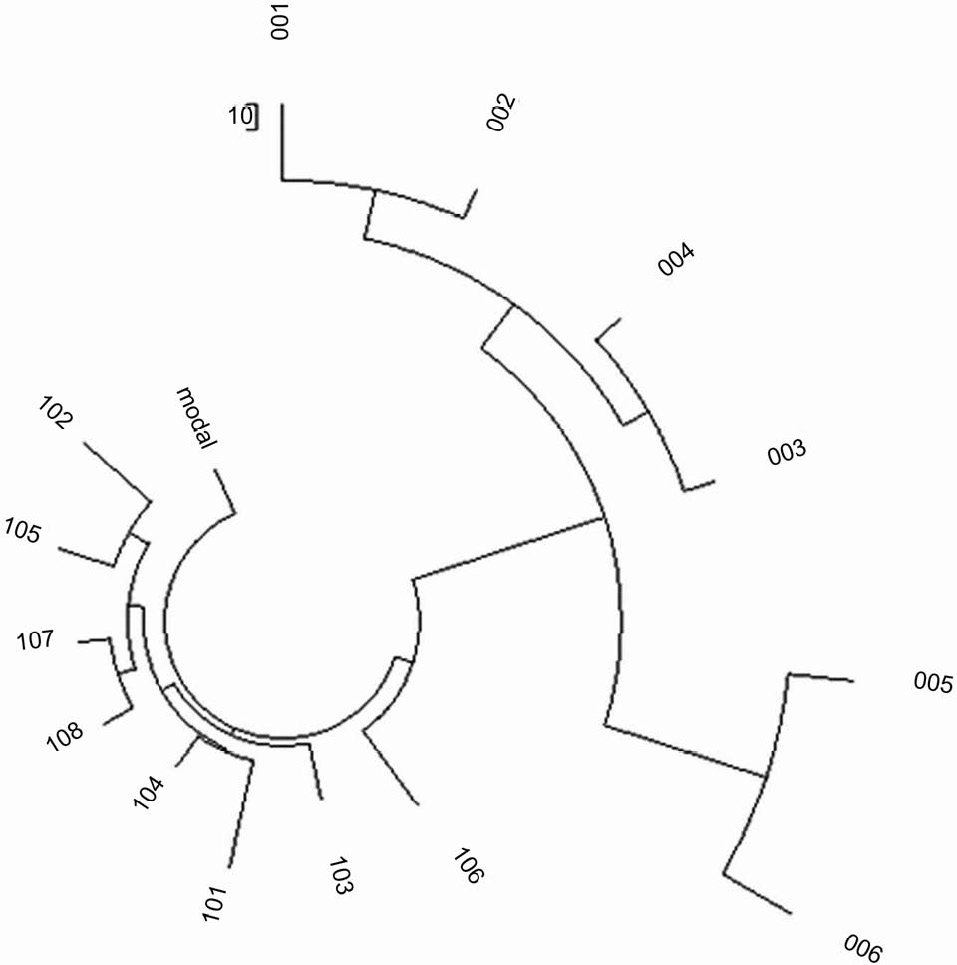
Рисунок 3. Древо 25 25-маркерных гаплотипов для гаплотипа R1b1a1-M73:
Правая ветвь - пятеро Русских, трое Казахов, один Китаец; нижние ветви - четыре Узбека, два Таджика,
один Турок, один из Люксембурга; левая ветвь - два Англичанина, два Итальянца, один Француз и один
Испанец.
Данные взяты из проекта FTDNA, в котором много гаплотипов доступны в 67-маркерном формате:
http://www.familytreedna.com/public/R1b1b1/default.aspx?section=yresults |
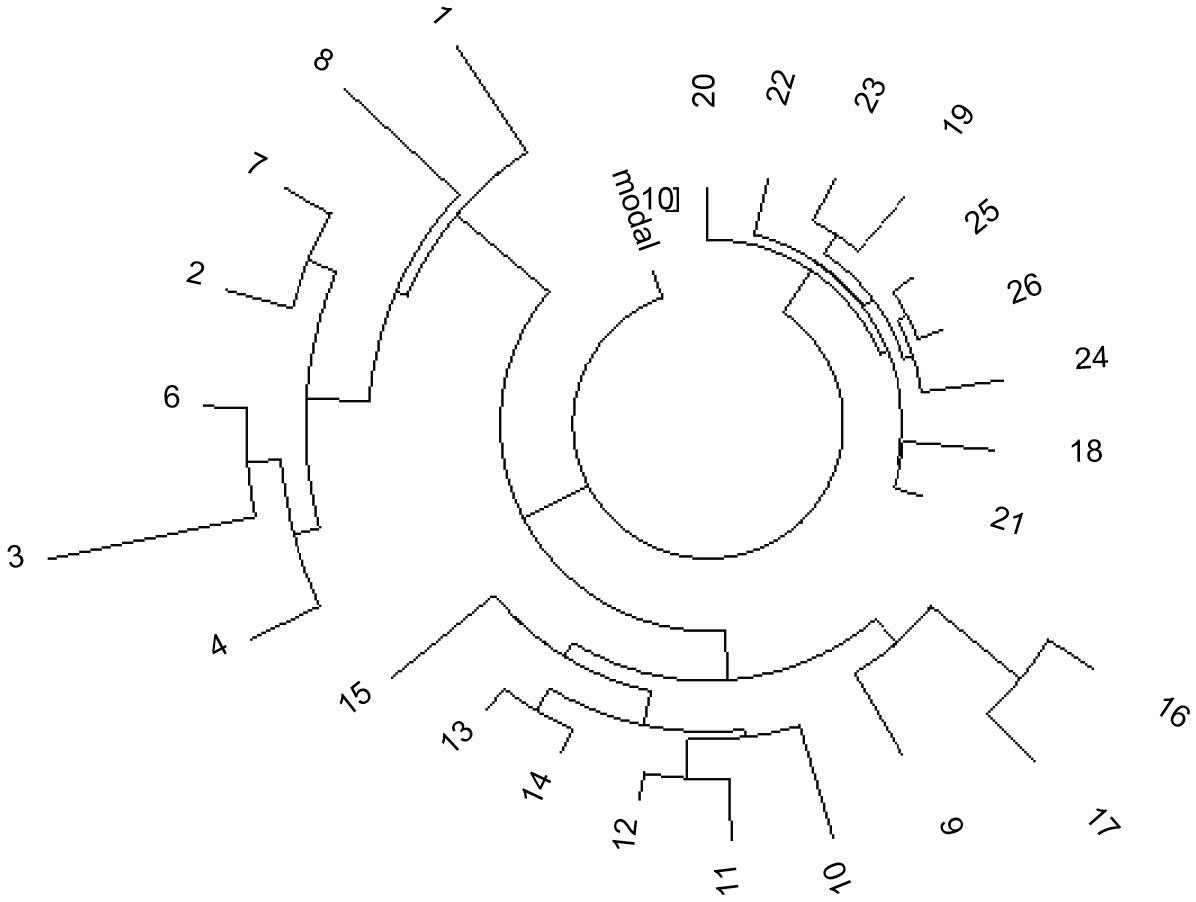
Тесная ветвь из 8-ми гаплотипов близко к “стволу древа на Рисунке 2 и правая ветвь из
9-гаплотипов на
Рисунке 3 имеют следующий одинаковый базовый гаплотип:
13 19 14 11 13 13 12 12 13 14 13 30 - 17 9 9 11 11 23 15 19 33 12 15 15 16 - 10 10 19 25 15 16 16 17
30 36 12 10 - 11 8 16 16 8 10 10 8 11 10 12 23 23 16 10 12 12 16 8 12 24 21 13 12 11 13 11 11 12 11
(M73, ветвь)
Замечательной особенностью этой базового гаплотипа (и каждого гаплотипа в обеих ветвях) является
второй аллель DYS390 = 19. В 25-маркерном формате, ветви содержат соответственно 15 и 17 мутаций,
что в обоих случаях дает те же 1075±280 лет до общего предка.
Следует отметить, что аналогичная серия из десяти гаплотипов R-M73 (с DYS 390 = 19) была получена
у населения Башкирии (Myres et al., 2010) недалеко от границы между Европой и Азией, по обе стороны
Уральских гор. Их базовый гаплотип был
13 19 14 10 Х Х Х 12 13 14 13 29 - 10
(последний маркер DYS461), и все сто аллелей в десяти гаплотипах имеццли только три мутации, что
дает 3/10/.018 = 17 поколений, или 425±250 лет до общего предка. Однако, как будет показано ниже,
общий предок этих и других R-M73 гаплотипов жил более 10 тысяч лет назад. В этих гаплотипах мы по
существу видим “верхушку айсберга”; они чрезвычайно далеки друг от друга и должны быть очень далеки
от их общего предка во времени. Похоже, что “база” айсберга поговорки исчезла, не оставив своих
древних гаплотипов. Иными словами, мы здесь видим ряд современных гаплотипов, которые либо прибыли
из других мест несколькими веками ранее, или прошли через тяжкие узкие демографические заторы.
Башкирские (аналогичные или слегка мутированные) гаплотипы R-M73 были выявлены в соседних
областях среди населения Татар и Мари (там же). В том же исследовании, R-M73 были обнаружены также
среди Кабардино-Балкарцев, Менгрелов, Турков, у всех Тюркоговорящих или Уралоговорящих,
принадлежщих к семейству Алтайских языков. Еще одно название, Черемисы, для Марийского народа в
соседнем языке Коми означает “люди с Востока”. Их базовый гаплотип
13 22 14 11 Х Х Х 12 13 14 13 32 - 10
отличается от базового Башкирского гаплотипа L23 на 7 мутаций (!) в десяти маркерах, помещая их
общего предка на 9700 лн (Клесов, 2011a), а если включить гаплотипы с DYS390 = 19, их общий предок
перемещается глубже во времени по крайней мере к 16.000 лет назад.
Возвращаясь к древу на Рисунке 3, нижняя ветвь в древе имеет следующий базовый гаплотип
(когда продлен до 67 маркеров)
13 22 14 11 13 17 12 12 12 13 13 30 - 15 9 9 11 11 23 15 20 30 12 15 15 17 - 11 10 19 24 15 14 17 17
35 39 12 10 - 11 8 16 16 8 10 10 8 10 10 12 23 23 15 10 12 12 18 9 12 25 20 13 12 11 13 11 11 12 12
(M73, ветвь)
и ее общий предок жил 3525±600 лн. Это соответствует правой ветви на Рисунке 2.
Левая боковая ветвь на Рисунке 3 имеет следующие базовый гаплотип:
12 25 14 11 13 14 12 12 12 14 13 29 - 16 9 10 11 11 22 15 20 32 12 15 16 17 - 11 10 19 25 15 17 17
18 34 36 14 10 - 11 8 16 16 8 11 10 8 11 10 12 22 23 17 10 12 12 15 8 13 22 20 14 12 11 14 11 11 12
12 (M73, ветвь)
и его общий предок жил 3050±600 лн.
Все три ветви довольно “молодые”, однако все они очень далеки друг от друга. Их попарные
мутационные различия 38, 38 и 42, что производит разделение по времени между их общими предками
11.500 - 13.200 лет мутационной эволюции их гаплотипов (мы это называем “боковым временем” между
двумя базовыми гаплотипами). Все три базовых гаплотипа M73 коллективно отличаются друг от друга на
62 мутаций, которые отводят их общего предка на ~7750 лн. Однако их мутационные разница становится
еще более выраженной по сравнению с базовым гаплотипом субклада R1b1a2-P312, предковым многим
Европейским субкладам гаплотипов R1b1a2. Для указанных выше M73 базовых гаплотипов она составляет
48, 35 и 42 мутаций, или до 16.000 лет мутационных различий между двумя базовыми гаплотипами. Это
определяет их общего предка как 10,600 лн.
Другая серия 26 от 12-маркерный M73 гаплотипы были перечислены в (Малярчук et al, 2011)., И были
собраны среди сибирских популяций: шорцев, телеутов, Калмыков, хакасов, Туве и Алтайцев. Эти
гаплотипы разделились на две ветви. Первый из них подходит именно на первые 67-маркерный базового с
имеющихся 12-маркерныйв, а второй отклоняется от базового гаплотипа соответствующих правая ветвь на
Рисунке 2 в двух аллелей (отмечен):
13 19 14 11 13 13 X X 13 14 13 30 - 15 10
13 22 14 11 13 16
X X 13
13 13 30 - 15 10
Первые 12-маркерный базовый гаплотип - смешанный Сибирско-Урало-Кавказского населения (считая их
текущих носителей — Башкиры, Калмыки, Тувинцы, Татары, Кабардинцы, Российцы и Алтайцы). Все 11
гаплотипов этой ветви коллективно насчитывают 20 мутаций, что дает 20/11/.024 = 76 → 83 условных
поколений, или 2075±510 лет до общего предка. Логарифмический метод дает [ln(11/2)]/.024 = 71 → 77
поколений, или 1925 лет, что почти то же (среди 11 гаплотипов в наборе данных 2 гаплотипа идентичны
по своим базовым гаплотипам. 0,024 - это константа скорости мутация по 12-маркерному гаплотипу). Те
же самые, но в 67-маркерном формате, Среднеазиатские гаплотипы M73 показали 1075±280 лет до общего
предка. Таким образом, Сибирско-Урало-Кавказский набор несколько “старше” по сравнению со
Среднеазиатским набором данных.
Второй 12-маркерный базовый гаплотип в основном был из Сибирской популяции (Шорцы, Телеуты,
Хакасы), а также единичные Мари, Турк и Татарин. Все 15 гаплотипов этой ветви насчитывают 22
мутации, что дает 22/15/.024 = 61 → 65 условных поколений, или 1625±380 лет до общего предка.
Логарифмический метод дает [ln(15/5)]/.024 → 48 поколений, или 1200±580 лет, что в рамках
погрешности то же самое. Однако, 67 -маркерный базовый гаплотип показал 3525±600 лет до общего
предка. Таким образом, тут Среднеазиатская ветвь M73 “старее” по сравнению с Сибирской. В целом,
Сибирские и Среднеазиатские гаплотипы были примерно в той же самой оценке “возраста".
Два 12-маркерных гаплотипа отличаются на 8 мутаций, которые отделяют друг от друга их двух общих
предков на 8/.024 = 333 → 492 поколений, или 12.300 лет. Это дает (12.300 +2075 +1625)/2 = 7985 лн
до их общего предка. Можно видеть, что данные, полученные с 67- и 12-маркерными гаплотипами субклада
M73 из Сибири и Средней Азии являются достаточно воспроизводимыми, и указывает на общего предка
гаплотипа M73 из Центральной Азии почти 8000 лн. Следует иметь в виду, что эти данные относятся к
живущим в настоящее время потомкам тех общих предков. Мы также должны помнить, что 67-маркерный M73
Центральноазиатский гаплотип показывает расстояние от Европейских гаплотипов R1b1a2 до 48 мутаций,
или до 16.000 лет мутационных различий между двумя базовыми гаплотипами. Это относит их (R-M73 в
Центральной Азии и R-M269 в Европе) общего предка на 10.600 лн.
90
Субклад гаплотипа R1b1-L278/R1b1*
Самый древний субклад R1b зарегистрированный Проектом R1b1 (xP297); xP297 не включает субклады
M73 и M269, рассмотренные в предыдущем разделе этой статьи. Все 68 гаплотипов проекта, доступные в
67-маркерном формате, отображены на древе Рисунок 4. Дерево гаплотипов включает в себя
субклад R1b1*, парагруппу от R1b1-L278 (см. показанную выше диаграмму R1b), V88, ее нижестоящий
субклад V69, и серию гаплотипов R1b1a2-M269, которые попали в проект, по-видимому, по ошибке (
однако, она полезна в древе как сравнительная ветвь).
Подробный анализ древа был выполнен в (Клесов, 2011a). Гаплотипы парагруппы R1b1* проекта - либо
Евреи-Ашкенази (из Венгрии, России, Беларуси) с их очень недавними общим предками всего несколько
веков назад, или очень разнобойной географии - из Италии, Пуэрто-Рико, Германии, Армении, Турции.
| R1b1c4-V88 |
R1b1c4-V69 |
R1b1a2 | R1b1a1-M73 |
| 1-11 | 12 | 13 | 14 | 15 | 16 | 17-19 | 20-22 |
24-39 | 40 | 41 | 42-47 |
| |
| Ашк. | Инд. | Арм. | Узб. | Ирак | Бахр. | Герм. |
Ашк. | Сефард. | СаудАр | Суд. | - |
Европа | Каз./Уйг./Узб./Тадж/Тув.. |
Формально, тройная ветвь справа на Рисунке 4 (гаплотипы 1 - 10) имеет общего предка,
жившего около 5800 лн. Предок не несет никакого определенного исторического значения, судя по их
составу, за исключением промежутка времени соответствуетвующему субкладу Р-L23 на Кавказе и в
Анатолии (Клесов, 2010d); конечно, R-L23 найдена не только там. Те разнобойные гаплотипы могли быть
принесены их предками из Центральной Азии вместе с R-M269, с последующим древом 67-маркерных
гаплотипов из 68 гаплотипов субкладов R1b1* (1 - 17), R1b1c-V88 (18 - 41), R1b1c4-V88- V69 (42 -
47), R1b1a2 (48 - 68). Дерево было составлено по данным FTDNA Проекта
http://www.familytreedna.com/public/R1b1Asterisk/default.aspx?section=yresults.
| Субклад R1b1-L278/R1b1* (1 - 17) проливает свет на одну из самых интересных и загадочных страниц
в истории Европы. Еврейское Ашкеназ/Ashkenaz (мн. Ашкеназим/Ashkenazim) относится к скифам
Аскузам/Askuz/Ishkuz, названию, которым именовались Тюркские Ас-кижи/As-kiji (Ас-люди) или
Ас-гузы/As-guz (Ас-племя), прозрачное обозначение Тюркского племени Асов ( Кит. Юэчжи/Yueji 月 支,
возможно составное Хуэ/Hui/Yu + чжоу/zhou 肉; Кит. наименование Суи/Хуэ/Юй/Ю обозначает Уйгуров и
предложительно обозначало Усунских/Wusun предков современных Уйсыней/Uisyns). Тем не менее, в
Библейском Иврите, Ашкеназ - это общее название для Тюркских племен независимо от племенной
принадлежности, и как таковая она была применена к Хазарам. Жизнь в Хазарском каганате была далека
от беззаботной, и религиозные потрясения были не лучше, чем политические, этнические или военные.
Первоначально Тенгрианское, население разделилось на чрезполосные религиозные фракции, после падения
Каганата и начала национально-религиозных преследований, потомки обращенных в Иудаизм оказались
отчужденными и беззащитными. Иудаизированные Тюрков в значительной степени рассеялись,
присоединившись к различным устойчивым Еврейским общинам, и образовали категорию по-Еврейски
называемую Ашкеназим (Тюрки) средневековой Европы. К счастью для них, Европа была разделена на
бесчисленное множество независимых и квази-независимых княжеств, и некоторые из них приветствовали
беженцев как денежно-кредитной и интеллектуальный капитал. Естественно, Ашкеназские мальчики несли
Y-ДНК их древних Тюркских предков, но их более близские предки вышли из смеси Тюркских племен
Хазарского Каганата, которые отпочковались тысячелетия назад, и накопили тысячелетия
независимого развития, примесей, и языковой и исторической эволюции.
Хазары были ответвлением Булгар, возможно разделявшим общие гены. Хазары, как и Булгары, были
отождествлены с древними племенами Асов. На генетическом древе, Хазары и Булгары скорее всего
образуют несколько отдельных парных ветвей. Отдельный главной ветвью Хазар были бы
Сувары/Савиры/Северяне, второе по величине большинство Каганата, которые имели свою собственную
историю, и вероятно различный генетический профиль. Специфически Булгарские субклады должны были
выжить среди Тюркских национальностей в Дунайской Болгарии; специфически Савирские субклады должны
иметь демографический пик в районе реки Десна, от Чернигова до Курска, в Украинском и Русском
населениях. Эти субклады должны также проявиться на древе исключительно Ашкенази ДНК, с
распределением между Булгарской и Савирской ветвями в соответствии с результатами Дунайской
Болгарии/Черниговщины.
На более глубоком уровне, скорее всего первоначальное появление Булгар и Хуннов в Восточной
Европе были событиями несвязанными, Хунны мигрировали с севера, из обласи Арала и верхнего
Итиля/Волги, а Булгары мигрировали с юга, из Бактрии/Тохаристана, или из современной восточной
Туркмении и юга Узбекистана. Гунно-Булгарами они стали уже в Восточной Европе. Наличие одинаковых
субкладов в южном Узбекистане и в спектре к востоку и западу от него поможет определить Булгарскую
ветвь, и таким образом поможет определить Савирскую ветвь. Савирская ветвь должны иметь повышенный
уровень остатков в районе Дагестана. Цифра 5800 лн является довольно значительной в этом отношении,
разделяя Булгарскую и Савирскую ветви за 2000 лет до появления Савиров/Субаров в Вавилонских записях
23-го в. до н.э. 28 веков спустя, когда Булгары и Савиры воссоединились, они говорили на языках
одной Огурской семьи, видимо, по-прежнему взаимопонятных.
Не все Ашкеназы произошли из Хазарских подданных. Некоторые могут иметь совершенно отличное
происхождение, как те, кто присоединился к Еврейским общинам из других Тюркских земель (С.-В.
Европа, Балканы, и т.д.), позжие выходцы из Кипчакского Ханства, Персидские Евреи прошедшие через
Кавказ, и тому подобное, или моли получили свое прозвище из-за связи с общинами Ашкенази. |
| Рисунок 4. Гаплотипы L23, со временем рассеяные по всей Европе |
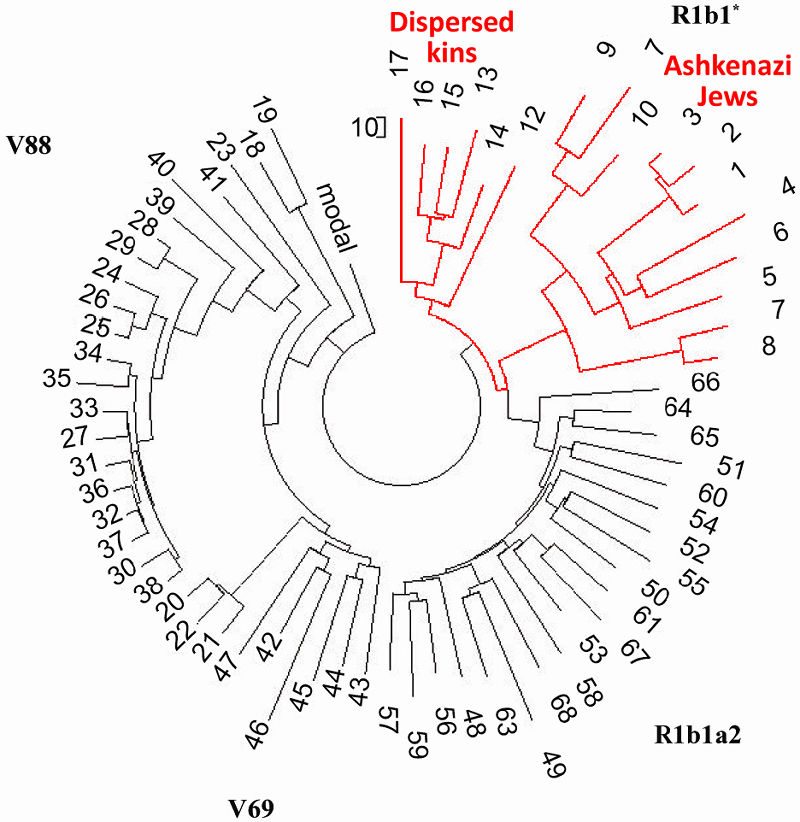
Следующая небольшой ветвь шести гаплотипов (12 - 17) включает гаплотипы из Центральной Азии
(Узбек), Армении, Ближнего Востока (Ирак, Бахрейн), Индии и Германии (последний сравнительно недалек
от Узбекского гаплотипа, в пределах 4900 лет мутационной эволюции). Индийский гаплотип имеет DYS390
= 18, что приближает его к Центральноазиатским гаплотипам с DYS390 = 19, рассмотренных выше. Другие
имеют DYS390 = 24 или 25. Их мутационные отклонения (без включения очень дальних гаплотипов) от
базового гаплотипа R1b1*
13 24 15 10 12 13/14 12 13 11/12 13 13 29 - 14 9 9 11 11 26 14/15 18 30 13/14 14 14/15 16 - 11 11 18
23 15 16 18 16 36 36 13 11 - 11 8 15 16 8 10 10 8 11 10 12 19 21 16 10 12 12 14/15 8 12 24 20/21 14
11 11 13 12 11 12 12 (R1b1*, ветвь)
в первых 25, 37 и 67 маркерах приводят к 238, 217, и 235 поколений от общего предка (без
поправки на обратные мутации), что составляет 230±11 поколений, достаточно воспроизводимое значение.
Самое надежное значение, для 67-маркерного формата, дает ~8400 лет до общего предка. Тем не менее,
Индийский гаплотип отличается от вышеприведенного базового гаплотипа на 62,5 мутаций (!), добавляя с
его DYS390 = 18 всего 6 мутаций в подсчет разницы. Это определяет отрезок времени(из имеющихся
гаплотипов) до общего предка R1b1* как 16.600 лет назад. Это, безусловно, указывает на
Центральную Азию, так как Европейско-Ближневосточные гаплотипы R1b1* достигают лишь “возраста” ~8400
лет.
Субклад R1b1c-V88 в Европе и Африке
На Рисунке 4 гаплотипы V88 занимают левую сторону древа . Опять же, часть их суб-ветвей
довольно "молода". Например, часть принадлежащая к ветви Евреев-Ашкенази (Россия, Украина, Венгрия,
Германия, Франция) с общим предком всего 350 лн. Гаплотипы Англии и Шотландии имеют общего предка
жившего 650 лн. Тем не менее, общим предком двух суб-ветвей (Евреи и острова) жил 6875 лн, на самом
дне субклада V88. Другая серия гаплотипов V88 определяет их путь из Саудовской Аравии (6225 лн)
через Испанию к Еврейской общине (1525 лн, в середине 1-го тысячелетия н.э.).
| R1b1c4-V88 |
| 1-11 | 12 | 13 | 14 | 15 | 16 | 17-19 | 20-22 |
24-39 | 40 | 41 |
| Ashk. | Ind. | Arm. | Uzb. | Iraq | Bahr. | Germ. |
Ashk. | Sefard. | SauAr | Sud. |
Базовый гаплотип ветви V88 на Рисунке 4 выглядит следующим образом:
13 24 16 10 13 14 12 12 12 14 13 29 - 16/17 9 10 11 12 26 14 19 29 12 12 15 15 - 11 12 21 23 15 15
18 19 33 35/36 12 11 - 12 8 15 16 8 10 10 8 9 11 12 22 23 15 11 12 12 15 8 12/13 23 20 13 12 11 13
11 12 12 12 (V88)
и его носителя жили 6575±700 лн.
Расширенная серия 72-х 11-маркерных гаплотипов V88 из Африки была предоставлена в (Cruciani et
al., 2010). Они разделились на две очень явные ветви и разнобойные серии гаплотипов (Клесов, 2010b).
Одна ветвь из 37 гаплотипов имеет следующий базовый гаплотип (в формате - 413a,b 460, 461, GATA A10,
YCAIIa,b после первых стандартных 12-ти маркеров, некоторые из них отсутствуют):
13 X 15 11 X X X X 12 X X X - 23 21 11 10 13 21 23
Все вместе содержат 111 мутаций, что дает 111/37/.02 = 150 → 176 условных поколений, или 4400±610
лет до общего предка. Этот Африканский базовый гаплотип V88 не укладывается ни с одним Евразийским
базовым гаплотипом V88; однако, он достаточно хорошо укладывается с базовым гаплотипом R1b1c4-V69
(см. следующий раздел), чей общий предок жил 4300±600 лн. Статья (Cruciani et al, 2010) предложила
еще один субклад обозначенный R1b1a4, с 29 гаплотипами. Их базовый гаплотип совпадает с показанным
выше; все 29 гаплотипов содержат 76 мутаций, и дает 76/29/.02 = 131 → 150 поколений, или 3750±570
лн, что находится в пределах погрешности указанной даты.
91
Другая ветвь V88 Африканских гаплотипов V88 - недавняя с общим предком 750±290 лн. Все 72
Африканские гаплотипы имеют те же базовый гаплотип, как показано выше с ее 201 мутациями, что дает
201/72/.020 = 140 → 163 поколений, или 4075±500 лн, в пределах допустимой погрешности для основной
ветви V88 , однако первый подсчет более точный, так как он был рассчитан на определенную ветвь.
Эта дата, 4300±600 лн, хорошо вписывается в миграционный путь носителей R1b с Ближнего Востока на
запад вдоль побережья Средиземного моря до Атлантического океана между 5500 и 4800 лн (см. ниже).
Кажется, что племя R1b-V88-V69 возможно разделилось, изменив направление миграции на юг, в Камерун и
Чад, где носители R1b-V88 живут сегодня.
Субклад R1b1c4-V69
Это субклад представлен на Рисунке 4 (No 42-47) шестью
гаплотипами в нижней части древа, с базовым гаплотипом
13 23 15 11 13 15 12 12 13 13 13 29 - 16/17 9 10 11 12 27 14 19 29 12 12 14 15 - 11 12 21 23 15 16
17 19 33 36/37 12 12 - 12 8 15 16 8 10 10 8 10 11 12 21 24 15 11 12 12 14 8 12/13 22 20 13 12 11 13
11 12 12 13 (V69)
Все они коллективно имеют 106 мутаций от базового гаплотипа, что дает 106/6/.12 = 147 → 172
поколений, или 4300±600 лет от общего предка. Это согласуется с “возрастом” вышестоящего субклада
V88, с возрастом ~6575 лн. Базовый гаплотип V69 отличается на 16 мутаций от базового гаплотипа V88
(см. выше, некоторые мутации дробные), что разделяет их общих предков на 16/.12 = 133 → 154
поколений, или 3850 лет, и помещает их общего предка на (3850 + 6575 + 4300)/2 = 7400±800 лн. Это в
пределах погрешности с результатом 6575±700 лн для вышестоящего субклада V88.
Анализ показывает, что R-V88 является довольно “молодым” субкладом по сравнению со всей R1b, и
R-V69 является “моложе”. Возможно, он возник при миграции R1b на общем маршруте к Ближнему Востоку,
например, на северо-востоке или востоке от Каспийского моря, или в западной части Ирана.
Субклад R1b1a2-M269
Последняя ветвь на древе на Рисунке 4 в нижней правой части (No 48,
56-57, 59) имеет базовый гаплотип
13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 - 11 11 19 23 15 15 18
17 37 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12
12 (M269, смесь)
Это точно базовый гаплотип субкладов R-P312 и R-L21. Они являются типичными и широко
распространенными Европейскими гаплотипами. Все 20 гаплотипов ветви содержат 365 мутаций, что дает
365/20/.12 = 152 → 179 поколений, или 4475±505 лет до общего предка. Это, опять же, является
типичным “возрастом” для смеси гаплотипов субклады R-P312 и R-L21 и их нижестоящих субкладов.
Субклады M269 вместе с ее нижестоящими субкладами были проанализированы в многочисленных
публикациях (Клесов, 2008a, 2009d, 2010c, 2010d, 2010e, 2011a, 2011b). Короче говоря, это субклад
возник около 7000 лн в Центральной Азии или в восточной части Восточно-Европейской равнины - его
немедленно нижестоящий субклад R-L23 (возник ~6000 лн) широко распространен среди Башкир на Южном
Урале, в Севернм Казахстане и прилегающие регионах, и их можно увидеть в России. Он мигрировал на юг
до Кавказа и за его пределы, в Анатолию и на Ближний Восток. Почти все основные Европейские и
Ближневосточные субклады гаплогруппы R1b (в настоящее время более чем 80 субкладов) являются
производными от субклада R1b1a2-M269.
Следует отметить здесь, что два братских субклада, R1a и R1b, мигрировали из Центральной Азии на
запад по двум совершенно различным маршрутам. В то время как группы R1a двигались вдоль южного
маршрута из области Алтая через Гималаи, Индостан, Иранское нагорье, Анатолию и остальную Малую Азию
до Балкан (Клесов и Рожанский, 2012), R1b двигались по северному маршруту, из той же области через
Южный Урал, Среднюю Азию, Северный Казахстан, Средняю Волгу, Кавказ, а затем разделились между южным
и западным направлениям. Эта миграционная модель также объясняет, почему носители R1a, но не R1b
найдены в Восточных Гималаях (Kang et al, 2011; См. также Клесов и Рожанский, 2012).
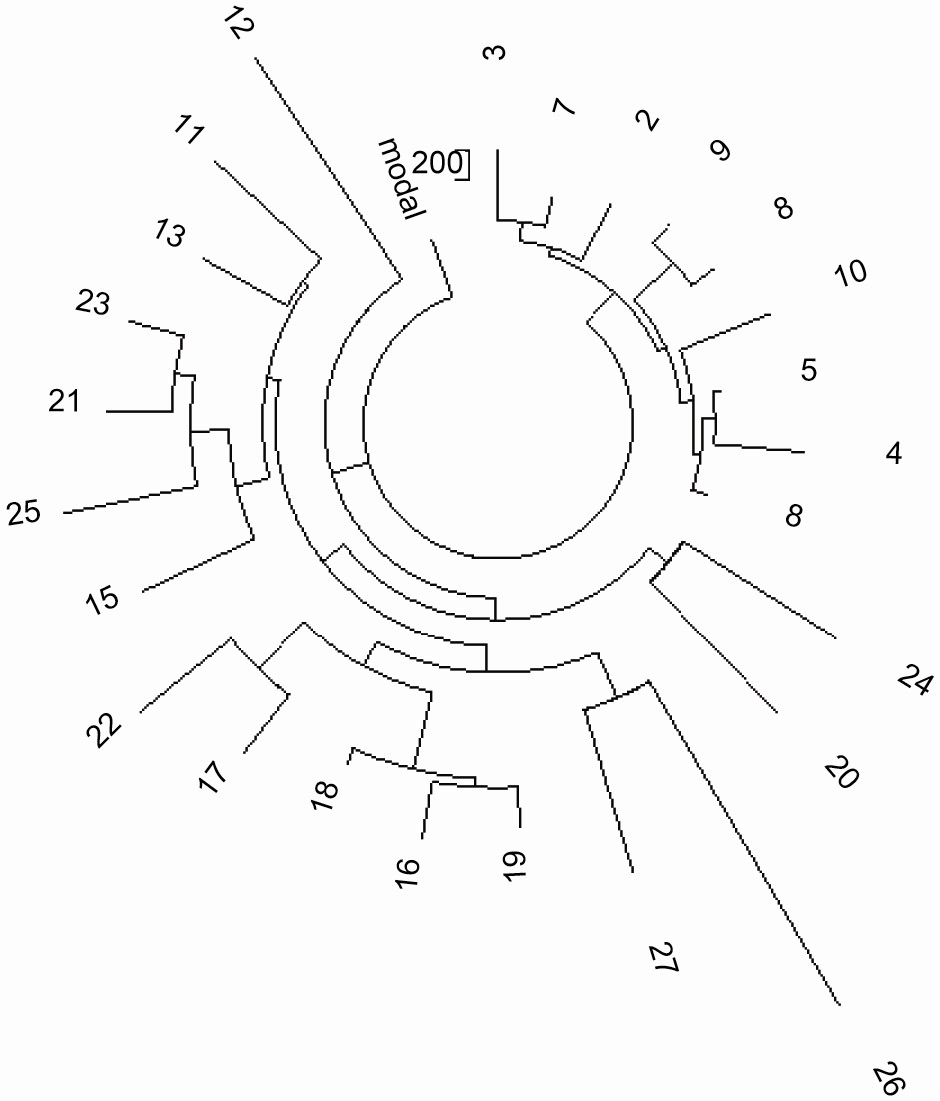
На Рисунках 5 и 6 показаны соответственно 37- и 67-маркерные древа субкладов гаплотипа
R1b1a2*-M269. Древа содержат две различные ветви, одна довольно сложная и, очевидно, “старая”, с
базовым гаплотипом
12 25 14 11 11 14 11 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 16 17 - 10 11 19 23 15 15 19
17 35 38 12 12 - 11 9 15 16 8 10 10 8 11 10 12 23 23 16 10 12 12 15 8 13 22 20 13 12 11 13 11 11 12
12 (M269)
и другая, плотная, плоская, “младшя”, и в значительной степени Еврейская ветвь, с базовым
гаплотипом
12 24 14 10 11 14 11 12 12 13 14 29 - 17 9 10 11 11 25 15 19 30 15 15 16
16 -
11 10 19 23 17 16 17 17 38 39 13
12 - 11 9 15 16 8 10 10 8 11 10 12 21 23 16 10 12 12
14 8
12 22 20 13 12 11 13 10 11 12 12 (M269, Еврейская)
Рисунок 5. 37-маркерное древо 25-ти гаплотипов для гаплотипов R1b1a2*-M269
Старшая (~6225 лн) ветвь слева и внизу включает гаплотипы из Турции, Армении, Италии, Франции,
Англии. Младшая (1000 лн) ветвь в правом верхнем углу включает Украину, Беларусь, Польшу и
Казахстан.
http://www.familytreedna.com/public/ht35new/default.aspx |
 92
92
Рисунок 6. 67-маркерное древо 25-ти гаплотипов для гаплотипов R1b1a2*-M269
того же набора данных как показано на Рисунке 5, однако, с меньшим количеством доступныx
гаплотипов |
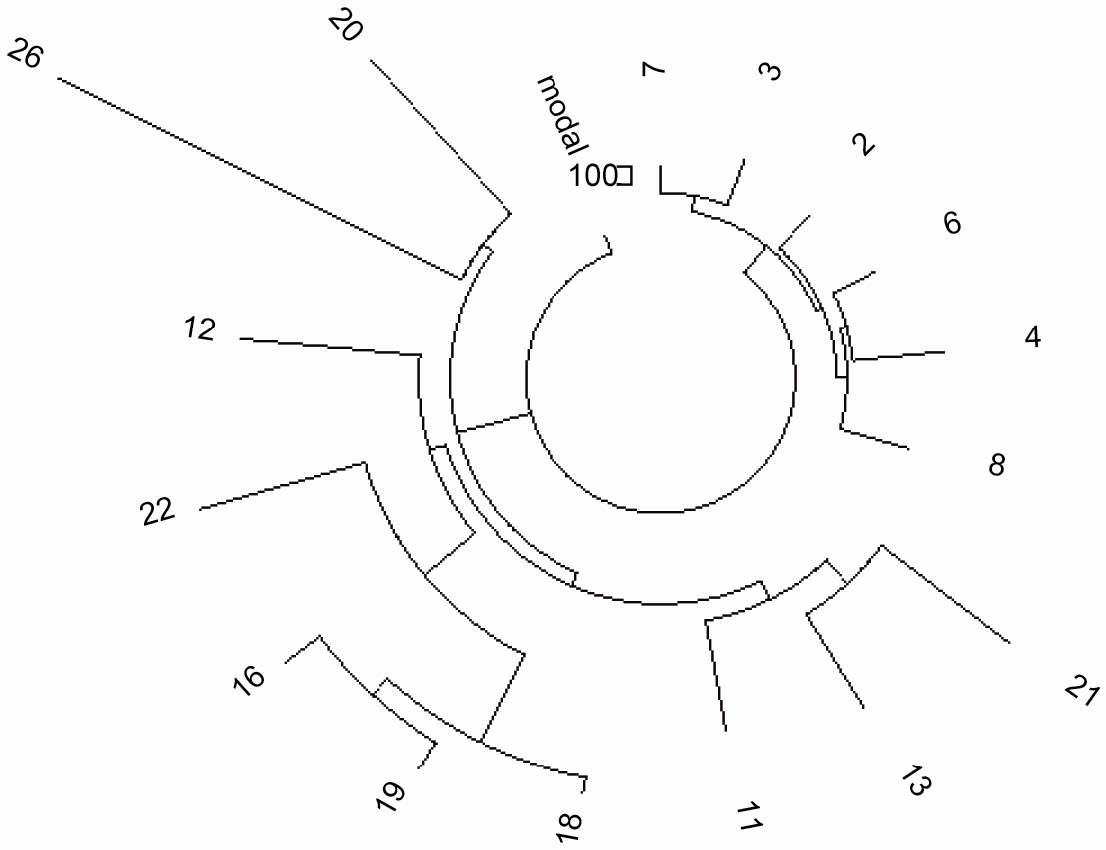
Мутационные различиия между двумя типами выделены жирным шрифтом. Младшая ветвь имеет 26 мутаций
во всех шести 67-маркерных гаплотипах, что дает 26/6/.12 = 36 → 37 условных поколений, или 925±200
лет до общего предка. 37-маркерная ветвь содержит 31 мутацию девяти гаплотипов, что дает
31/9/.09 = 38 → 40 поколений, или 1000±200 лет, по сути то же самое. Это Еврейское население
(“младшее”, Рис. 5), показаное с 14-ю 37-маркерными гаплотипами, было
проанализировано ранее (Клесов, 2008b) и определено, что их общий предок жил 1100±250 лн; базовый
гаплотип в точности такой же, как и у “молодой “ветви” данной выше.
| Время демографического затора Еврейских гаплотипов R1b1a2*-M269 также совпадает с дисперсией
Ашкенази после падения Хазарского Каганата. |
Старшая ветвь имеет общего предка, жившего 6200±900 лн. Два базовых гаплотипа различаются на 22
мутации, которые разделяют их на 22/.12 = 183 → 224 поколений, или 5600 лет, а общий предок обеих
ветвей жил 6400±900 лн.
Это описание дает нам общую картину миграции Арбинов из Центральной Азии (~16000 лн) на запад в
течение следующих 10 - 9 тысяч лет, до примерно 7000 - 6000 лн.
Гаплогруппа R1b (в основном R1b1a2-L23) среди Башкир, и на Кавказе, в Анатолии,
на Ближнем Востоке
По-видимому, R-L23 возник в восточной части Восточно-Европейской равнины, где Европа встречается
с Азией, ~6200 лн, и мигрировал на Кавказ и далее на юг, в Анатолию и на Ближний Восток. Другая
ветвь L23 ответвилась на запад, в Европу, около 4500 лн.
Самое восточное население с преобладающим субкладом R-L23 - это Башкиры, Тюркоязычный народ,
которые в значительной степени живет по обе стороны Уральских гор и в Северном Казахстане. Частота
гаплогруппы R1b достигает 84% у Пермских Башкир, 81% среди Баймакских Башкир, и более низкие цифры у
других Башкирских племен (Лобов, 2009). Были опубликованы 29 Башкирских 10-маркерных гаплотипов
субклада R-L23 (Myres et al., 2010), и их базовый гаплотип такой
12 24 14 10 Х Х Х 12 12 13 13 30 - 10.
Это типичный, хотя и немного мутировавший гаплотип L23 с его характерным первой аллелью DYS393 =
12. Из этих 29, 26 гаплотипов были идентичны, как показано выше, и вся Башкирская ветвь L23 имеет
общего предка, жившего только 575±175 лн. Тем не менее, этот базовый гаплотип отличается от
Европейского R-L23 базового гаплотипа
12 24 14 10 Х Х Х 12 12 13 14 29 - 11
на три мутации, которые отделяют друг от друга эти два базовых гаплотипа на 3/.018 = 167 → 200
условных поколений — 5000 лет, и помещают их общего предка на 5500 лн. Другие Башкирские гаплотипы
принадлежат R-M269 (один гаплотип), R-M73 (10 гаплотипов) и R-U152 (8 гаплотипов). В последней серии
гаплотипов они все идентичны друг другу, и таким образом выходят от самого недавнего общего предка,
который безусловно имел Европейское происхождение [U152 возник в Европе 4125±450 лн (Клесов,
2011b)].
| Мария Гимбутас датировала Третью Курганную Волну, вторжение Причерноморских “Ямных” Курганцев в
Центральную и Восточную Европу ок. 3000 г. до н.э. ~5000 лн |
Гаплотипы R1b в Myres et al., 2010, Дополнительная Таблица S3, для няций с сильным Тюркским
присутствием
Без этнической идентификации образцов, этническая атрибуция невозможна (напр. Турция)
Для многонациональных народов, как Башкиры, Кумыки, Турция, Иран (Персия), выборки крайне
недостаточны |
Башкиры
| L23 |
14 |
12 |
13 |
17 |
24 |
10 |
13 |
12 |
12 |
10 |
N=21 |
44% |
| M73, M478 |
14 |
12 |
14 |
16 |
19 |
10 |
13 |
13 |
13 |
10 |
8 |
17% |
| U152 |
15 |
12 |
13 |
17 |
24 |
11 |
13 |
13 |
12 |
11 |
8 |
17% |
| L23 |
14 |
12 |
13 |
17 |
24 |
10 |
13 |
12 |
11 |
10 |
2 |
4% |
| L23 |
14 |
12 |
13 |
17 |
24 |
10 |
13 |
12 |
13 |
10 |
2 |
4% |
| L23 |
14 |
12 |
13 |
17 |
23 |
10 |
13 |
12 |
12 |
10 |
1 |
2% |
| L23 |
14 |
12 |
13 |
17 |
25 |
10 |
13 |
12 |
12 |
10 |
1 |
2% |
| L23 |
14 |
12 |
13 |
16 |
23 |
11 |
13 |
12 |
13 |
10 |
1 |
2% |
| L23 |
14 |
12 |
13 |
17 |
24 |
10 |
13 |
13 |
12 |
10 |
1 |
2% |
| L23 |
15 |
12 |
13 |
17 |
24 |
10 |
13 |
12 |
11 |
10 |
1 |
2% |
| M73, M478 |
14 |
12 |
12 |
16 |
19 |
10 |
13 |
13 |
13 |
10 |
1 |
2% |
| M73, M478 |
14 |
12 |
14 |
16 |
19 |
10 |
13 |
13 |
12 |
10 |
1 |
2% |
Балкары
| M73, M478 |
14 |
12 |
14 |
19 |
22 |
11 |
13 |
13 |
12 |
10 |
N=3 |
30% |
| M73, M478 |
14 |
12 |
14 |
19 |
23 |
11 |
13 |
13 |
13 |
10 |
2 |
20% |
| M73, M478 |
14 |
12 |
14 |
18 |
22 |
11 |
13 |
13 |
12 |
10 |
2 |
20% |
| L23 |
14 |
13 |
13 |
17 |
25 |
11 |
13 |
12 |
11 |
10 |
1 |
10% |
| M73, M478 |
14 |
12 |
15 |
19 |
22 |
11 |
13 |
13 |
13 |
10 |
1 |
10% |
| M73, M478 |
14 |
12 |
14 |
18 |
22 |
11 |
13 |
13 |
13 |
10 |
1 |
10% |
| M73, M478 |
14 |
14 |
13 |
19 |
22 |
11 |
13 |
13 |
13 |
10 |
1 |
10% |
Кумыки
| L23 |
14 |
12 |
14 |
16 |
24 |
11 |
13 |
12 |
14 |
10 |
N=1 |
33% |
| L23 |
14 |
12 |
13 |
15 |
24 |
11 |
15 |
12 |
12 |
10 |
1 |
33% |
| L23 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
12 |
12 |
10 |
1 |
33% |
Кайтаки (Каи-таги/Kayi-tag/Kaitak/Kayitag), R1b1b2, (Балановский et al.,
2012)
| M269 |
13 |
28 |
23 |
16 |
14 |
11 |
14 |
17 |
15 |
13 |
19 |
13 |
10 |
13 |
12 |
13 |
23 |
12 |
12 |
| M269 |
13 |
29 |
24 |
17 |
14 |
11 |
14 |
16 |
15 |
12 |
19 |
12 |
11 |
13 |
12 |
12 |
23 |
12 |
12 |
Татары
| M73, M478 |
14 |
12 |
14 |
16 |
19 |
10 |
13 |
13 |
12 |
10 |
N=1 |
50% |
| M73, M478 |
14 |
12 |
13 |
17 |
22 |
11 |
13 |
13 |
13 |
10 |
1 |
50% |
Даргинцы
| L23 |
14 |
12 |
14 |
17 |
23 |
11 |
12 |
12 |
12 |
10 |
N=1 |
50% |
| L23 |
15 |
12 |
13 |
16 |
23 |
11 |
13 |
12 |
13 |
11 |
1 |
50% |
Авары
| L23 |
14 |
12 |
12 |
16 |
24 |
11 |
13 |
12 |
13 |
10 |
N=1 |
100% |
Менгрелы
| M73, M478 |
14 |
12 |
14 |
19 |
21 |
11 |
13 |
12 |
13 |
10 |
N=1 |
50% |
| L23 |
14 |
12 |
14 |
17 |
24 |
11 |
13 |
12 |
12 |
10 |
1 |
50% |
С. Осетинцы (Дигоры или Ардоны)
| L23 |
14 |
12 |
12 |
18 |
24 |
11 |
13 |
12 |
12 |
11 |
N=1 |
50% |
| L23 |
14 |
12 |
12 |
17 |
24 |
11 |
13 |
12 |
12 |
11 |
1 |
50% |
Табасаранцы
| L23 |
14 |
13 |
13 |
16 |
23 |
11 |
13 |
12 |
12 |
10 |
N=2 |
33% |
| L23 |
14 |
12 |
12 |
16 |
23 |
10 |
13 |
12 |
12 |
11 |
1 |
17% |
| L23 |
14 |
12 |
13 |
16 |
23 |
11 |
13 |
12 |
14 |
10 |
1 |
17% |
| L23 |
14 |
12 |
13 |
16 |
23 |
10 |
13 |
12 |
13 |
10 |
1 |
17% |
| M269 |
15 |
12 |
13 |
16 |
25 |
10 |
13 |
12 |
11 |
10 |
1 |
17% |
Иран
| M269 |
14 |
12 |
14 |
16 |
24 |
11 |
12 |
13 |
12 |
10 |
N=1 |
100% |
Мари
| M73, M478 |
14 |
12 |
13 |
17 |
23 |
10 |
13 |
13 |
14 |
10 |
N=1 |
50% |
| M73, M478 |
14 |
12 |
14 |
16 |
19 |
10 |
13 |
13 |
13 |
10 |
1 |
57% |
Пакистан
| L23 |
14 |
13 |
13 |
16 |
24 |
10 |
13 |
12 |
12 |
10 |
N=1 |
20% |
| L23 |
14 |
12 |
13 |
17 |
24 |
11 |
13 |
14 |
12 |
11 |
1 |
20% |
| L23 |
14 |
12 |
13 |
15 |
25 |
11 |
14 |
12 |
13 |
9 |
1 |
20% |
| L23 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
12 |
12 |
10 |
1 |
20% |
| L23 |
14 |
12 |
15 |
15 |
24 |
11 |
14 |
12 |
12 |
11 |
1 |
20% |
|
Турция
| L23 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
12 |
12 |
10 |
N=6 |
8% |
| L23 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
12 |
13 |
10 |
4 |
5% |
| M269 |
14 |
12 |
13 |
16 |
25 |
11 |
13 |
12 |
11 |
10 |
3 |
4% |
| L23 |
14 |
12 |
14 |
16 |
24 |
11 |
13 |
13 |
11 |
11 |
2 |
3% |
| L23 |
14 |
12 |
13 |
16 |
23 |
10 |
13 |
12 |
12 |
10 |
2 |
3% |
| L23 |
14 |
12 |
13 |
15 |
24 |
11 |
14 |
12 |
12 |
10 |
2 |
3% |
| L23 |
14 |
12 |
13 |
15 |
24 |
11 |
14 |
12 |
13 |
10 |
2 |
3% |
| L23 |
14 |
12 |
13 |
16 |
24 |
10 |
13 |
12 |
12 |
12 |
2 |
3% |
| L23 |
14 |
12 |
12 |
16 |
24 |
11 |
13 |
13 |
14 |
10 |
2 |
3% |
| L23 |
15 |
12 |
12 |
15 |
24 |
10 |
13 |
12 |
11 |
11 |
1 |
1% |
| L23 |
13 |
12 |
13 |
16 |
25 |
11 |
13 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
12 |
15 |
24 |
10 |
14 |
12 |
12 |
11 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
13 |
12 |
11 |
1 |
1% |
| L23 |
14 |
12 |
13 |
15 |
24 |
11 |
15 |
12 |
13 |
10 |
1 |
1% |
| L23 |
14 |
13 |
14 |
16 |
23 |
10 |
13 |
12 |
11 |
10 |
1 |
1% |
| L23 |
15 |
12 |
13 |
16 |
24 |
11 |
13 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
21 |
10 |
13 |
12 |
12 |
10 |
1 |
1% |
| L23 |
15 |
12 |
13 |
16 |
21 |
10 |
13 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
17 |
25 |
10 |
13 |
12 |
12 |
10 |
1 |
1% |
| L23 |
15 |
12 |
12 |
15 |
23 |
10 |
14 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
12 |
15 |
24 |
10 |
14 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
15 |
24 |
10 |
15 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
24 |
10 |
13 |
12 |
13 |
11 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
12 |
11 |
10 |
1 |
1% |
| L23 |
13 |
12 |
13 |
18 |
25 |
11 |
13 |
12 |
11 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
15 |
24 |
10 |
14 |
12 |
13 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
12 |
13 |
11 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
25 |
11 |
13 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
24 |
10 |
13 |
12 |
13 |
10 |
1 |
1% |
| L23 |
13 |
12 |
13 |
17 |
24 |
11 |
13 |
12 |
14 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
24 |
10 |
14 |
12 |
13 |
10 |
1 |
1% |
| L23 |
15 |
12 |
13 |
16 |
23 |
11 |
13 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
24 |
12 |
13 |
13 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
15 |
23 |
10 |
14 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
24 |
10 |
12 |
12 |
12 |
11 |
1 |
1% |
| L23 |
14 |
12 |
12 |
16 |
25 |
11 |
13 |
12 |
13 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
13 |
12 |
10 |
1 |
1% |
| L23 |
15 |
12 |
14 |
15 |
25 |
10 |
12 |
12 |
11 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
15 |
23 |
11 |
11 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
14 |
17 |
25 |
10 |
13 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
15 |
24 |
10 |
14 |
12 |
11 |
11 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
24 |
10 |
13 |
12 |
12 |
10 |
1 |
1% |
| L23 |
14 |
12 |
13 |
16 |
23 |
11 |
13 |
12 |
11 |
11 |
1 |
1% |
| L23 |
14 |
12 |
14 |
17 |
23 |
10 |
13 |
12 |
12 |
11 |
1 |
1% |
| L23 |
13 |
12 |
13 |
17 |
25 |
11 |
13 |
12 |
12 |
10 |
1 |
1% |
| M269 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
12 |
12 |
10 |
1 |
1% |
| M269 |
14 |
12 |
13 |
16 |
25 |
10 |
13 |
12 |
13 |
10 |
1 |
1% |
| M269 |
14 |
12 |
13 |
18 |
24 |
10 |
13 |
12 |
11 |
10 |
1 |
1% |
| M269 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
13 |
11 |
10 |
1 |
1% |
| M269 |
14 |
12 |
14 |
16 |
24 |
11 |
13 |
12 |
11 |
10 |
1 |
1% |
| M269 |
14 |
12 |
14 |
16 |
24 |
11 |
12 |
12 |
11 |
10 |
1 |
1% |
| M269 |
14 |
12 |
12 |
15 |
24 |
10 |
14 |
12 |
12 |
11 |
1 |
1% |
| M412 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
13 |
12 |
12 |
1 |
1% |
| M73, M478 |
14 |
12 |
13 |
17 |
23 |
11 |
13 |
13 |
14 |
10 |
1 |
1% |
| S116 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
13 |
12 |
12 |
1 |
1% |
| S116 |
15 |
12 |
12 |
17 |
24 |
10 |
13 |
13 |
13 |
11 |
1 |
1% |
| U106 |
14 |
12 |
13 |
15 |
23 |
11 |
13 |
13 |
12 |
11 |
1 |
1% |
| U106 |
14 |
12 |
13 |
16 |
25 |
11 |
14 |
13 |
11 |
12 |
1 |
1% |
| U152 |
14 |
12 |
12 |
16 |
24 |
11 |
13 |
14 |
11 |
10 |
1 |
1% |
| U152 |
14 |
12 |
13 |
16 |
24 |
11 |
13 |
14 |
12 |
12 |
1 |
1% |
| U152 |
14 |
12 |
13 |
16 |
24 |
11 |
15 |
13 |
12 |
12 |
1 |
1% |
|
Таким образом, мы видим сегодняшнее отражение древних миграций Арбинов из Центральной Азии на
запад, по-видимому из области Ю.Сибири, через Южный Урал и далее на Восточно-Европейскую равнину, а
затем на Кавказ. Почти все гаплотипов R1b1a2 в области Кавказа относятся к субкладу L23 (с
характерным DYS393 = 12). В недавней работе (Балановский et al., 2012) были перечислены 90
Кавказских гаплотипов гаплогруппы R1b, и за исключением пяти гаплотипов R1b* и сравнительно
“молодой” Абхазский ветви (Рис. 7), в наборе данных 79 из 81 гаплотипов (97,5%) были субклад
L23 (Примечание: цитируемая работа не просчитывала древья гаплотипов, ни проанализировала гаплотипы
представленной здесь методике).
Тот же шаблон наблюдается и с Армянскими гаплотипами R1b, и с большинством Анатолийских R1b
гаплотипов (Клесов, 2010c, 2011c). Полученный из расширенного набора данных гаплотипов со всего мира
(древо Рис. 8) 67-маркерный базовый гаплотип R-L23 выглядит следующим образом:
12 24 14 11 11 14 12 12 12 13 13 29 - 16
9 10 11 11 25 15 19 29 15 15 16 17 - 11 11
19 23 16
15 17 17 36 37 12 12
- 11 9 15 16 8 10 10 8 10 11 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (L23)
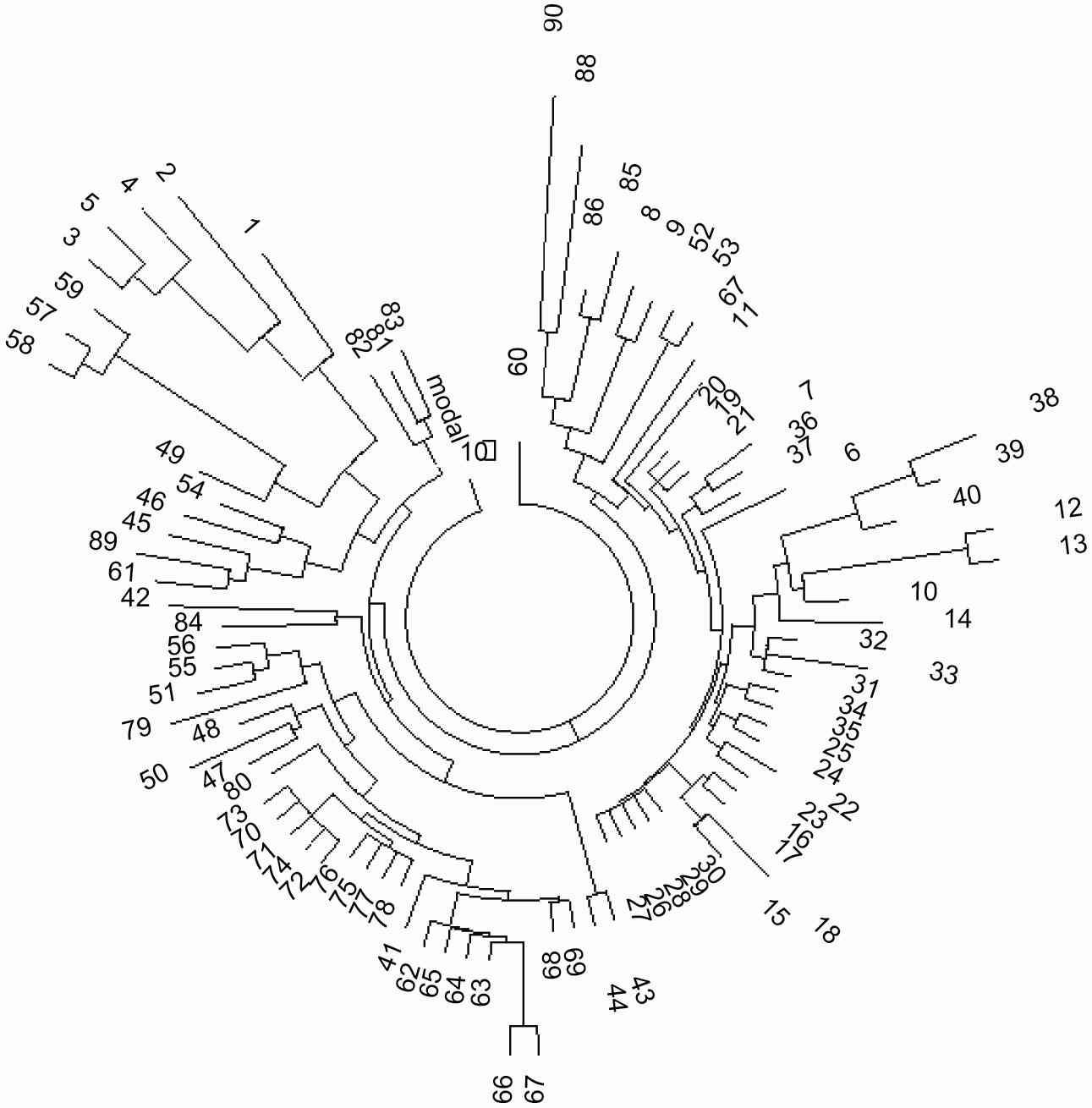
Короткий Кавказский базовый гаплотип L23 на Рисунке 7 точно подходит именно к
вышеназванному гаплотипу (соответствующие аллели выделены жирным шрифтом). Общий предок субклада L23
жил ~6200 лн (Клесов, 2010d, 2011a). 81 Кавказских гаплотипов L23 с 425 мутациаями от их базового
гаплотипа дают 425/81/.035 = 150 → 176 условных поколений, или 4400±490 лет до общего предка.
“Молодая” дата (по сравнению с “возрастом” L23 около 6200 лн) может быть объяснена подробным
рассмотрением расширенной серии из 107-ми гаплотипов R-L23, перечисленных в Проекте FTDNA (см.
надпись на Рисунке 8). Древо на
Рисунке 8 распадается на две части. Слева находятся 38 гаплотипов, с базой
12 24 14 11 12 15 12 12 12 13 13 29 - 16 9 10 11 11 25 15 19 30 15 15 16 18 - 11 11 19 23 16 16 18
17 36 37 12 12 - 11 9 15 16 8 10 10 8 11 11 12 23 23 15 10 12 12 16 8 12 22 20 13 12 11 13 11 11 12
12 (L23, ветвь)
93
Рисунок 7. 19-маркерное древо 90 гаплотипов гаплотипа R1b на Кавказе (Лезгины, Осетины,
Аварцы, Анхазы, Черкесы, Нахи (Чеченцы), Даргинцы, Каитаги
(Кайтаки))
Гаплотипы 1 - 5 (вверху слева) представляют R1b* субклад,
и гаплотипы 49, 57 - 59 (вверху слева) представляют собой “молодую” Абхазскую ветвь. Гаплотипы
перечислены в (Балановский et al., 2012) |
Справа находятся 69 гаплотипов, с базой
12 24 14 11 11 14 12 12 12 13 13 29 - 16 9 10 11 11 25 15 19 29 15 15 16
17 - 11 11 19 23 15 16 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 11 12
23 23
16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (L23, ветвь)
Более 70% гаплотипов Армян и Турков из серии относятся ко второй и большей ветви, а также все
восемь Иракцев и все пять Иранцев серии. Присутствие R-L23 у Иранцев может быть результатом диффузии
субклада из Анатолии на восток, или миграция Арбинов могла быть на юг из Восточно-Европейской
равнины к востоку от Каспийского моря.
Обе ветви спустились со своего предкового базового гаплотипа R-L23, и разошлись на 9 мутаций
(выделены жирным шрифтом). Эти 9 мутаций накапливались в течении 9/.12 = 75 → 81 условных поколений,
или 2025 “боковых” лет. Первая ветвь раскололась 4600±490 лн, вторая 4200±440 лн. Таким образом, их
общий предок жил (4600 + 4200 + 2025)/2 = 5400±800 лн. Это вписывается в погрешность времени, когда
жил общий предок субклада R-L23 (~6200 лн). Базовый гаплотип расширенных, 111-маркерных гаплотипов
из той же выборки, и для небольшой ветви, выглядит следующим образом:
12 24 14 11 12 15
12 12 12 13 13 29 - 16 9 10 11 11 25 15 19 30 15 15 16
18 - 11 11 19 23 16 16 18 17 36 37 12 12 - 11 9 15 16 8 10 10 8 11 11 12
23 23
15 10 12 12 16 8 12 22 20 13 12 11 13 11 11 12 12 - 36
15 9 15
12 25
26 19 12 11 13 12 10 9 12 12 10 11 10 30 12 13 24 13 10 10 19 15 19 13 24 17
12 15 24 12 23 18 10 14 17 9 11 11 (L23, ветвь)
Он отличается на 16 мутаций от более крупной ветви (7 мутаций добавлены расширением 68 - 111),
которые отделяют ветвь на 16/.198 = 82 → 90 поколений, или 2250 лет, и общий предок R-L23 жил (4600
+ 4200 + 2250)/2 = 5525±700 лн - аналогично 5400±800 лн полученным выше, и иллюстрирует
согласованность расчетов.
Похоже, что Кавказские гаплотипы R-L23 с общим предком 4400±490 лн принадлежат одной из ветвей
древа на Рисунке 8. Гораздо меньшая Кавказская выборка R1b, проанализированная ранее (Клесов,
2008а), привела к похожему времени - 4650±700 лн, как недавний, больший набор коротких гаплотипов
(Балановский et al, 2012.). Кавказские гаплотипы R-L23 возможно испытали демографическую горловину
около 5000 лн.
Недавно были опубликованы в (Herrera et al., 2011) 120 17-маркерных Армянских гаплотипов. Древо
гаплотипов из тех гаплотипов показано на Рисунке 9. Оно состоит из трех примерно равных (по
количеству, или “весу”) ветвей. Пять гаплотипов M269 мини-ветви
(which ones?) почти равны друг другу, что свидетельствуя о
только двух мутациях на 85 аллелей. Их общий предок жил 300±210 лн.
| Ок. 1600 гг. Османская империя и Сефевидская Персия разделили Армению, северо-западная Армения
была аннексирована Османской империей; это кажется отражено в молодой мини-ветви из пяти гаплотипов
M269. |
Остальные 115 гаплотипов субклада L23 насчитывают 784 мутаций от их базового гаплотипа
12 24 14 11 11 14 XX 13 13 13 29 - 17 15 19 12 15 12 23
(правая сторона гаплотипа соответствует DYS 458, 437, 448, GATA H4, DYS 456, 438, 635).
94
Это дает 784/115/.034 = 201 → 250 условных поколений, или 6250±660 лет от общего предка. Это
опять же типичный отрезок времени до общего предка субклада R-L23, и находится в согласии с
“возрастом” ~7000 лн его вышестоящего субклада R-M269.
Тот же базовый гаплотип, как показано в предыдущем пункте, был найден в наборе из 238 Армянских
шести-маркерных гаплотипов R1b опубликованых ранее (Weale et al., 2001) и проанализированы в
(Клесов, 2008a). Он включал гаплотипы из шести регионов Армении, Карабаха, Ирана и других областей
Армянской диаспоры. Он может быть представлен в виде
12 24 14 11 X X X 12 X X 13 X.
Средний “возраст” общего предка гаплотипов R1b во всех шести регионах был 5750±1500 лет, что
соответствует другим оценкам субклада R-L23.
Происхождение Армян от Балканских Фригийцев хорошо сновано на классических источниках; Армянский
- это агглютинативный язык; Фригийский язык не было взаимно понятен с языком Троянцев и с
окружающими Анатолийскими языками. Этнографически, мы знаем Фригийский колпак, совпадающий со
Скифским башлыком и традиционными башлыками Тюркских народов; мы также знаем Фригийский выстрел,
совпадающий со Скифским выстрелом и легендарными выстрелами Тюркских всадников; Фригийский король
Гордион был похоронен в традиционном Скифском и Тюркском кургане, окруженном многими другими
курганами, что указывает на Курганную экспансию на Балканы. Таким образом, Курганный маркер R1b у
Армян соответствует их происхождению по классическим источникам, однако, Армяне имеют Тюркские
примеси от Античности до Позднего Средневековья, и их древо R1b должно иметь многочисленные ветви
образованные в этот период, которые должны иметь параллельные ветви среди многочисленных
отдельных Тюркских народах.
 |
Рассматривая другие популяции с довольно высокой долей гаплотипов R-L23, которые в свою очередь
могут быть следами древней миграции Арбинов, может быть упомянута Россия с 37% известных гаплотипов
R1b имеющих DYS393 = 12 (Roewer et al., 2008; Клесов, 2010c с ссылками) и базовый гаплотип
12 24 14 11 11 14 X X 12 13 13 29/30 - 16 15 19 (последние три аллели DYS 458, 437, 448),
такой же, как R-L23 показаный выше, и с общим предком Русского R1b 6775±830 лн (Клесов, 2009d);
Анатолии и Ливана с их
12 24 14 11 X X X 12 13 13 29 и
12 24 14 10 X X X 12 13 13 29 (мутировавшая DYS391 11 → 10 отмечена)
и с соответствующими общими предками 6000±820 лн (Анатолия) и 5200±670 лн (Ливан) [Клесов, 2010c;
гаплотипы были перечислены в Cinnioglu et al, 2004, и Zalloua et al, 2008].
Среди Еврейских гаплотипов R1b (большинство из них не было типировано на субклады несколько лет
назад, так как типирование еще не производилось), одна ветвь удивительно напоминает гаплотипы R-L23,
и ее 37-маркерный базовый гаплотип был следующим (Клесов, 2008b):
12 24 14 10 11 14 12 12 12 13 14 29 - 17 9 10 11 11 25 15 19 29 15 15 16 17 - 11 11 19 23 16 16 18
17 37 38 12 12 (L23, Еврейский)
В данном конкретном случае набор данных содержал 42 37-маркерных гаплотипов, которые в
совокупности имели 620 мутаций, что дает 620/42/.09 = 164 → 196 поколений, или 4900±530 лет от
общего предка. Базовый гаплотип отличается на 9 мутаций от “молодого” Еврейского базового гаплотипа
указанного выше в первых 37-ми маркерах, что верроятно делает гаплотип с 4900 лн предочным. Самый
старый определенный Еврейский базовый гаплотип R1b датирован 5400±500 лн (Клесов, 2008b). Он,
безусловно, появился на Ближнем Востоке в Шумерскую эпоху.
| Так как определено, что наличие базового гаплотипа R1b является результатом миграции, следующим
шагом будет расссмотреть, какие миграции могла принести его. В доисторический период, ДНК-генеалогия
может предложить миграции и эффект основателя, но письменный период может предложить другие
возможности. Наличие большой массы Тюркских религиозных терминов (Адам, Ева, Эдем) в Библии
указывает на период до-Скифской, но пост-Неолитической миграции, и период 2500-2000 г. до н.э.,
отвечает этим требованиям (Туруки, Гутии, Субары, Куманы, Канги). Потом была миграция Скифского
периода, и классические источники помещают Скифского гуру и других актеров в Ближневосточную среду.
В Новой Эре, Хуннские и Савирские походы разыгрывается на Ближневосточной сцене, и с Арабским
завоеванием массы Кипчаков и других Тюркских рабов были ввезены на Ближний Восток, и это все до
систематического заселения первоначально Центральноазиатскими и Сибирскими племенами Ближнего
Востока, начиная с 9 в. н.э. Теоретически, ничто не мешает всем этим волнам принести на Ближний
Восток линии с общими предками намного старше, чем приход волн, а тогда даты можно обнаружить только
в случаях эффекта основателя. |
95
Шумеры, вероятные Носители гаплотипов R1b1a2, и их потомки Ассирийцы
Ассирийцы являются одной из старейших сохранившихся групп, происходящих, как полагают, от
исторических Шумеров. Среди Ассирийцев R1b является основной гаплогруппой, достигая 40% исследуемых
популяциий; гаплогруппа J занимает второе место с 11%, а остальные находятся в единичных процентах
(Lashgary et al, 2011). Такой высокий процент гаплогруппы R1b очень необычен в Южной Месопотамии. По
Ассирийскому FTDNA Проекту (см. Приложение) половина носителей R1b принадлежит к субкладу
R-L23 с DYS393 = 12, и все девять 12-маркерных гаплотипов имеют 35 мутаций, что дает 35/9/.02 = 194
→ 240 условных поколений , или 6000±1200 лет до общего предка. Только 6 гаплотипов были доступны в
25-маркерном формате, и они имеют 48 мутаций, что дает 48/6/.046 = 174 → 211 поколений, или 5275±930
лет. Это действительно Шумерские времена (напр. Kramer, 1971). Большинство носителей этих гаплотипов
сейчас живут в Ираке и Турции. Другая половина Ассирийских гаплотипов в Проекте, в основном из
Ирана, имеют слегка мутированный “классический” Европейский базовый гаплотип R1b1a2 (с DYS464 = 15
15 17 17), и общего предка 850±360 лн, рассчитанное по первым 12-ти маркерам, и 1100±280 лн по
первым 37-и маркерам. Эти данные по R1b очевидно указывают на относительно недавние события в
Ассирийском населении, принесенные из Европы.
| Не существует известного массового движения Ассирийцев из Европы 1000 лет назад, но в самом деле
1000 лет назад было одно из основных передвижений людей, связаное с Тюркским захватом Ближнего
Востока и начиная с Месопотамии, и оно может быть разумным источником относительно недавних событий
и слегка мутированного “классического Европейского” базового гаплотипа R1b1a2 принесенного из
Центральной Азии. |
Рисунок 9. 17-маркерное древо с 120 гаплотипами гаплогруппы R-L23, за исключением пяти
гаплотипов субклада M269 (номера 1 - 5)
Древо составлено на основе данных (Herrera et al., 2011) |
Субклад R-L23 четко прослеживается от Восточно-Европейской равнины на юг через Кавказ, где он
преобладает среди гаплотипов R1b, и через Анатолию, где он очень выражен; вниз до Южной Месопотамии,
где Шумеры жили между 6000 и 4500 лн ( Kramer, 1971). Так как время до общих предков тех гаплотипов
R-L23 около 6000 - 5000 лн, вполне вероятно, что эти общие предки жили среди Шумеров и их предков
(на Кавказе и в Анатолии). Это может быть дополнительным аспектом для лингвистов, некоторые из
которых считают Шумерский остатком подгруппы Дене-Кавказской языкового суперсемейства (напр.
Bengtson, 1997).
| Многочисленные авторы связывают Шумерский с Тюркскими языками, и эндоним Шумера Канг с
Центрально-Азиатским народом Кангар; исследования показывают, что Шумерский имеет множество Тюркских
заимствований вперемешку с Шумерским субстратом (см.
Тюркский-Шумерский для примера
работ и библиографии); исследования также находят многочисленные Венгерские изоглоссы, потенциально
также происходящие из Тюркской лексики. Таким образом, есть совпадающие лингвистические,
ономастические и биологические доказательства Шумеро-Тюркских исторических связей. Ссылка на
Дене-Кавказское языковое суперсемейство (суперсемейство включающее Северо-Кавказские языки,
Сино-Тибетские языки, и Кетский как Енисейский язык) также значительна, исследования показывают, что
агглютинативные Америндийские языки имеют слишком много системных соответствий с Тюркскими
агглютинативными языками чтобы предположить случайное совпадение. Эта значительность особенно
важная, так как временное разделение между Американскими и Евразийскими предковами оценивается в
15.000 лет, что указывает на уникальную стабильность агглютинативных языков как языкового явления
(см.
Содержание Америнд-Тюркские для примера
работ и библиографии). |
Миграция Арбинов из Понтийских Степей, Малой Азии и Ближнего Востока на запад в
Европу
В описаниях выше, мы оставили носителей гаплогруппы R1b на Кавказе/Восточно-Европейской
равнине/Причерноморских степях, в Анатолии и Ливане/Южной Месопотамии около 6000 - 5000 лн. Все три
области были для Арбинов транзитные, для продвижения далее на запад. Гаплотипы их нынешних потомков
служат указателями этих древних миграций.
| Их движение не могло быть ограничено “дальше на запад”, это наше эгоцентричное внимание
замораживает фокус нашего внимания в этой специфической манере. Похоже, что одинажды обследованные,
все известные области были зарегистрированы в долговрененной коллективной памяти, и формы миграций
скорее напоминают последовательные полеты бумеранга чем полеты стрелы, о чем свидетельствуют,
например, взаимонезависимые возвращения Асов и Каиев обратно на Кавказ. С различными
демографическими масштабами и различными временными параметрами, не все колебателные волны миграций
могут быть обнаружены. Исключительная, по сегодняшней демографической шкале, малочисленность
миграционных групп должна была вызвать эффект интерференции, с высокими гребнями и низкими
впадинами; литературные источники отражали гребни, в то время как демографическая генетика больше
видит впадины, генетический “шум” транзитных миграций. |
Набор данных по гаплотипам R1b1a2 на Балканах был опубликован в (Barac et al., 2003a, 2003b;.
Pericic et al, 2005, и частные контакты с М. Pericic). Он содержит ряд явно R-L23 гаплотипов с
базовым
12 24 14 11 11 15 Х Х Х 13 13 29
и общим предком 4050±890 лн (Клесов, 2010c). Другой ряд Балканских R1b1a2 имеет базу
13 24 14 11 11 11 Х Х Х 14 13 29,
с общим предком 4975±1300 лн (там же). Четыре мутации между ними (в том числе мутации с
множественными копиями, просчитывающиеся по-другому) помещают их общего предка на ~7000 лн ко
времени древних гаплотипов R-M269 Восточно-Европейской равнины (или далее на восток). Миграции на
запад из Понтийских степей также поддерживаются множественными археологическими свидетельствами,
которые включают даты третьего тысячелетия до нашей эры (напр. Mallory, 1989).
Древо гаплотипов показаное на Рисунке 8 дает некоторые подсказки относительно возможных
направлений этих древних миграций. 40% всех 107 гаплотипов древа относятся к Армянам и Туркам, из
них 70% гаплотипов на правой (большей) части древа. Верхняя левая ветвь в древе содержит гаплотипы
из России, Литвы, Польши, Хорватии и Ирландии.
96
Они все имеют одинаковую структуру мутаций, с базовым гаплотипом
12 24 14 11 11 14 12 12 12 12
13 29 - 16 9 10 11 11 25 15 19 31 14 15 16 18 (L23, в основном Восточная Европа)
который отличается от базового гаплотипа левой стороны 38-гаплотипной ветви на три мутации
(отмечены; некоторые аллели дробные). Это относит их общего предка вместе со всей ветвьа к 4600 лн.
Это означает, что вышеуказанная в основном Восточно-Европейская ветвь отделились от большой ветви на
Восточно-Европейской равнине в середине 3-го тысячелетия до нашей эры, и видимо мигрировала на
запад. Это один из маршрутов миграции (из нескольких уже описанных), объясняющий как гаплотипы R-L23
появились в те времена в Европе.
Другой миграционный путь проходил с Восточно-Европейской равнины на юг, как указывается другой
специфической ветвью на противоположной стороне (на 5 часов) на древе гаплотипов. Ветвь включает в
себя гаплотипы из России, Литвы, Армении (два гаплотипа), Турции (три гаплотипа), Сирии. Базовый
гаплотип выглядит следующим образом:
12 24 14 11 11 14 12 12 12 13
13 30 - 16 9 10 11 11 25 15 19 28 15 15 16
17 (L23, в основном на юге)
Между этими двумя базовыми гаплотипами ветвей, которые обе включают в себя и Русские и Литовские
гаплотипы, хотя и с принципиально иными историями, есть пять полных мутаций. Одна группа носителей
L23 пошла на запад в Европу, другая отправилась на юг, на Кавказ и на Ближний Восток. Они разделены
5/.046 = 109 → 122 поколений, или 3050 “боковыми” годами (в этом случае требуется произвести в
среднем пять мутаций в 25-маркерном гаплотипе). Другими словами, эти две ветви показывают вилку в
маршруте миграции R-L23 с востока Европейской равнины на запад и на юг.
| Это самое раздразнивающее наблюдение прошло без достойного комментария: 3050 "боковых" лет дают
1500 лет каждой ветви, что соответствует 500 г. н.э.; и 500-е гг. н.э. были самыми волнительным
моментом в истории Европы, когда Европейская Хуннская конфедерация раскололась, а ее народы
стратифицировались и мигрировали. Миграция на западе была относительно незначительной, указывая на
низкую подвижность пеших мигрантов, в то время как в северо-западные, восточные и южные миграции
были быстрыми и дальними. На следующем этапе истории мы сталкиваемся с внезапно появившимися без
объявления в южной Литве Хрватами/Хорватами/Chorbat Кангарами разделенными на Белых и Черных Хрватов
(Хорваты), где под Черными Хрватами мы можем разумно углядеть тех же внезапно появившимися с
Хорватами Сербов, которые были отмечены Птолемеем (ок. 130 н.э.) вблизи реки Итиль на западной
окраине Кангарского государства, их статус Черных Хрватов отражен в их наименовании Серб/Серф
(Крепостной) означающем зависимое племя; Акатирсы-Скифы/Древляне в Галиции были разгромлены
Булгарами и раскололись, большинство бежало на восток и на юг, и 1000 лет спустя в Турцию и Персию;
Савирские племена Хуннского Западного Крыла (Котургуры/Кутригуры ~ Köturgur/Kutriguri) отступили к
бассейну верхнего Дона и на Кавказ, внезапно создав без объявления свое господство на
северо-восточном Кавказе; и Северокавказские Булгары перешли в Паннонию и на северо-восточные
Балканы. Демографическая и политическая карта Восточной и Центральной Европы изменилась кардинально,
и ДНК-генеалогия позволяет взглянуть на соответствующие биологические следы. Примечательно, что в
500-м году нашей эры Славяне не известны в Восточной Европе, Восточная Европа делится между Финскими
(Уральскими) и Тюркскими народами, и соответственно мигранты R-L23 почти исключительно Тюркские
(лингвистически родственные Иранцам племена позже иавестных как Валахи и Балто-Славяне находятся в
Центральной Европе, в поясе север-юг, с вкраплениями Тюркских племен в средней степной зоне). |
Еще одна ветвь R-L23 включает в себя гаплотипы из Армении, Ливана, Болгарии, Италии, Франции,
Испании, Германии. Она расположена на 7 часов на древе гаплотипов на Рисунке 8. Их базовый
гаплотип
12 24 14 11 11 14 12 12 12 13 13 29 - 16 9 10 11 11 25 15 19 29 15 16 16 18
(L23, Юг и Юго-Запад)
отличается от предыдущей рассмотренной ветви лишь на 2,6 мутаций или 1550 “боковых” лет. Это
указывает на движение R-L23 с Ближнего Востока на запад, в Европу. Ее соседняя ветвь, с гаплотипами
из Турции (три гаплотипа), Греции и Албании имеет почти такой же базовый гаплотип, что и выше,
только с одной мутацией 12 → 13 в первой аллеле (DYS393) и 18 → 17 (17,3 в среднем) в последний
аллеле. По существу, это тот же маршрут миграции из Малой Азии в Европу. На самом деле, вся правая
большая ветвь на Рис. 8 состоит из суб-ветвей преимущественно с Армянскими и Турецкими (и
немного Иракских и Иранских) гаплотипами R-L23 с единичными включениями гаплотипов из Греции,
Германии, Нидерландов, Англии, Шотландии. Это отражает пути миграции Арбинов из Малой Азии и с
Ближнего Востока в Европу, главным образом между 4500 и 3500 ВР.
| Мария Гимбутас не обнаружила Курганной миграции "вторжения" с Ближнего Востока в любое
место Европы, и приписала Третью Курганную Волну, вторжение Причерноморских “Ямных” Курганцев в
Центральную и Восточную Европу ок. 3000 г. до н.э. ~5000 лн. Следуя нумерационной системе
М.Гимбутас, предполагаемая Курганная миграция с Ближнего Востока 2500-1500 до н.э. (4500-3500 лн)
будет именоваться Четвертой Курганной Волной, и следующие Пятой Курганной Волной (Киммерийцы),
Шестой Курганной Волной (Скифы), Седьмой Курганной Волной (Сарматы), Восьмой Курганной Волной
(Аланы), Девятой Курганной Волной (Готы), Десятой Курганной Волной (Хунны), Одиннадцатой Курганной
Волной (Булгары), Двенадцатой Курганной Волной (Авары), Тринадцатой Курганной Волной (Кангары/Бечены
~ Печенеги), Четырнадцатой Курганной Волной (Огузы), Пятнадцатой Курганной Волной (Кипчаки). Кипчаки
были последними Тенгрианскими Курганцами, мигрировавшими в Европу. Эта система нумерации опустит
Курганную миграцию с юго-запада ок. 3000 г. до н.э. (5000 лн). |
Можно ожидать, что некоторые миграции с Ближнего Востока в Европу связанны с этой мутацией DYS393
12 → 13 в субкладе R-L23. Однако другие, более поздние миграции, могли произойти с Европейского
континента на восток, и они принадлежат к недавним субкладам, таким как L51, L11, P312, U106, L21,
U152 и т.д. (Клесов, 2011b). Действительно, большинство Сардинских гаплотипов имеют DYS393 = 13, как
следующие базовые гаплотипы Сардинии (вычисляется из данных, приведенных в [Contu et al, 2008]):
13 24 14 11 11 14 X X X 13 X 29
13 24 14 10 11 14 X X X 13 X 29
13 24 14 10 11 15 X X X 13 X 29
Первый из них был представлен на древе серии одинаковых гаплотипов, очевидно происходящих от
очень недавнего общего предка (Клесов, 2008a, 2010c). Они ничем не отличаются от общеевропейских
субкладов R1b1a2-M269, таких как R-P312, R-L21, R-U152, R-L2, R-L20 и т.д. Второй имеет время до его
общего предка 3550±790 лн, третий произошел от общего предка, жившего 2900±620 лн. Общий предок их
всех жил 5025±630 лн, что подходит древней миграции Арбинов с Ближнего Востока в Европу по времени и
направлению (впрочем, DYS393 = 13 для них будет необычным), а из Иберии на север на континент, и
затем заселение Европы во всевозможных направлениях.
Один из наиболее распространенных базовых гаплотипов R1b в Сицилии выглядит следующим образом
(Gaetano Di et al, 2009.):
13 24 14 11 11 14 Х Х 12 13 13 29
что совпадает с одним из Сардинских базовых гаплотипов R1b упомянутых выше, и неотличимо от общего
субклада R1b в Европе. Для этого гаплотипа Сицилии промежуток времени 4550±1020 лет от общего предка
рассчитан в упомянутом исследовании. Тот же базовый гаплотип является наиболее распространенным в
Италии (Capelli et al., 2007), с временем до общего предка 4125±500 лет по линейному методу, и
4300±1160 лет по логарифмическому методу (Клесов, 2008a).
Существует еще один, очень важный миграционный путь Арбинов, который практически не включают
гаплотипы R-L23 (DYS393 = 12, только 5% среди Иберийских гаплотипов R1b, он соответствует случайной
мутации родительской DYS393 = 13). Это маршрут с Ближнего Востока на запад вдоль Северноафриканского
Средиземноморского побережья. R-L23 либо не представлен, или не выжил на этом пути. Кажется, что
носители R-V88 преодолели часть дороги, но однако они разделились и пошли на юг и обосновались в
Центральной Африке (в основном в Камеруне и Чаде), как упомянуто ранее. Миграция Арбинов с Ближнего
Востока до Атлантики, а затем через Гибралтарский пролив в Пиренеи состоялась от ~5500 - 5200 лн до
4800 лн, когда Арбины приземлились в Иберии (см. ниже подтверждающие ссылки на археологические
данные). Часть пути из Египта в Иберию могла быть по морю, подробности до сих пор пока не известны.
Есть некоторые исторические сообщения о прибытии Египетского военного флота в Иберию около 5000 лн,
есть некоторые якобы Египетские мумии, и фрагменты Египетских гробниц выставлялись в Королевской
Академии Истории в Мадриде, и в Таррагонском городском музее. Их статус, однако, довольно
расплывчатый. Но все таки представляется, что носители субкладов R1b, в основном R-M269 и вновь
образованные L51 и L11 (см. ниже) прибыли в Иберию, и это было началом археологической культуры
Колоколовидных Кубков.
До обсуждения Колоколовидных Кубков и истории Арбинов в Европе, стоит упомянуть еще одно довольно
расплывчатое свидетельство о путешествии R1b через древний Египет между 5500 и 5200 лн. Это касается
предполагаемого гаплотипа R1b фараона Тутанхамона.
97
Предполагаемый гаплотип R1b1a1-M269* фараона Тутанхамона и возможная его
история
Недавно Швейцарская компания iGENEA опубликовала заявленние о 16-маркерном гаплотипе фараона:
13 24 14 11 11 14 X X 10 13 13 30 - 16 14 19 10 15 12 (www.igenea.com/en/index.php?c=62).
Здесь первые 12 маркеров показаны в формате FTDNA, а остальные были 458 DYS, 437, 448, GATA H4, 456
DYS, 438. Это, очевидно, не типичный Европейский гаплотип R1b1a2, так как он имеет DYS439 = 10, а не
общеевропейское 12. Только около 0,5% гаплотипов R1b в Европе имеют DYS439 = 10. Наиболее вероятный
и наиболее тесно связанный базовый гаплотип - это R1b1a2-M269, как показано на
Рисунке 5, где те же маркеры, как и в гаплотипе фараона отмечены жирным шрифтом:
12 25 14 11 11 14 11 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 16 17 -
10
11 19 23 15 15 19 17 35 38 12 12 - 11 9 15 16 8 10 10 8 11 10 12 23 23 16 10 12 12
15 8 13 22 20 13 12 11 13 11 11 12 12 (M269)
Между двумя 16-маркерными гаплотипами есть 6,8 мутаций (некоторые аллели в гаплотипе M269
являются дробными), что переводится как 6.8/.0315 = 216 → 274 условных поколений, или 6850 лет между
ними. Так как Тутанхамон жил 3300 лн, и “возраст” базового гаплотипа R-M269 6200±900 лет, то общий
предок их двух жил (6850 + 6200 + 3300)/2 = 8175 лн. Это могло быть либо древний R-M269 предок
Тутанхамона из Центральной Азии, или соответствующий вышестоящий субклад.
Основным моментом здесь является то, что гаплотип фараона не какой-то ошибочно (или намеренно)
выбранный Европейский гаплотип R1b1a2; это архаичная гаплотип гаплогруппы R1b, вероятно из древнего
субклада R-M269.
Здесь следует отметить, что основателем Египетских фараонов был Нармер, его происхождение твердо
не известно, он был основателем Первой Династии, жил ок. тридцать второго века до н.э. - около 5200
лн. Эта дата соответствует времени миграции Арбинов вдоль побережья Средиземного моря в Северной
Африке. Не исключено, что носители гаплогруппы R1b на самом деле основали Королевский род в Древнем
Египте. Это не означает, что династическая линия не была прервана на протяжении Династий; однако она
постоянно возвращалась к Династиям, если происхождение некоторых фараонов действительно из линии
R1b.
Обзор Пути Миграции Арбинов, носителей гаплогруппы R1b, из Центральной Азии в
Европу
Общая карта маршрутов миграции R1b с самого “начала”, где “начало” точно не определено до сих пор
и может быть оценено между 16000 и 12000 лн, и до их прибытия на Пиренеи 4800 лн как будущих
Колоколовидных Кубкопв, показано на Рисунке 10. Их миграционные пути медленно формировались
начиная с Центральной Азии, видимо, с Алтая, в Южной Сибири, где были обнаружены некоторые очень
отличающиеся гаплотипы R1b. Как описано в первом разделе этой статьи, их огромная мутационная
разница с Европейскими и Ближневосточными гаплотипами R1b помещает их общего предка на ~16000 лн.
Рисунок 10. Карта расположения гаплотипов R1b настоящего исследовании, до их прибытия в
Иберию ~4800 лн
В целом путь миграции Арбинов, начался по-видимому в Южной Сибири, где сегодняшние гаплотипы R1b
Шорцев, Телеутов, Алтайцев, Уйгуров, Хакасов, Тувинцев (булавки справа) чрезвычайно отличаются от
гаплотипов в Европе и на Ближнем Востоке. Маршрут миграции проходил через области Средней Азии и
России, где в настоящее время живут Башкиры, Казахи, Узбеки, Марийцы, Татары, и Калмыки. Из области
Центральной России Арбины перешли на Кавказ и далее на юг в Анатолию, на Ближний Восток, в
современные Ирак, Ливан, а затем на запад вдоль Средиземного моря в Северной Африке до
Атлантического океана. На этом пути носители R1b-V88 отделились и углубились в Африку, где в
настоящее время живут их потомки в Камеруне и Чаде (Cruciani et al., 2010). Их миграционные пути в
Африку здесь воспроизводится из ссылки выше. Карта показывает места R1b до их прибытия на Пиренеи ~
4800 лн, откуда они двинулись на север как Колоколовидные Кубки и населили Европу между 4800 - 3000
лн и позже. |
Условные обозначения на Рисунке 10 описывает те народы Центральной Азии/Сибири. В той
области есть много Неолитических, Халколитических и Энеолитических археологических культур, таких
как Терсекская, Уральская, Суртандийская, Маханджарская, Иман-Бурлукская, Ботайская, Атбасарская,
Кельтеминарская, и другие археологические культуры Центральной Азии в современной России (напр.
Захаров, 2010), которые могут быть отнесены к Арбинам; однако, было бы преждевременным относить
любую из них к R1b или любой другой гаплогруппе. Такая задача является достаточно новой для
археологов. Заманчиво указать на Сероглазовскую, Хвалынскую, Самарскую, Средне-Волжскую и
прилегающие археологические культуры 12.000 - 5000 лн Европейского востока как наиболее вероятных
культур R1b. Однако мы не можем этого сделать по той же причине преждевременности, и было бы
безответственно предложить это в это время. То же самое можно сказать и о Срубной, Катакомбной и
соседних археологических культурах Центральной и Южной России, которые, видимо, были общими и для
R1b и для R1a носителей, хотя и в разные периоды времени. Люди R1b до 5000 лн, и люди R1a после 4500
лн спутали археологов, которые обозревали “разные корни” тех культур, распространяющихся в разных
направлениях и в разное время.
| Здесь мы сталкиваемся с интересным тезисом, что люди, которые разделились 10.000 лет раньше,
могли воссоединиться в одну археологическую культуру, как будто они только расстались внутри одного
поколения. В чисто терминах поколений, 10.000 лет составляют 400 поколений для каждого стержня
вилки. Эти 400 поколений отделены бесчисленными, и различными, технологическими инновациями,
генетическими и лингвистическими примесями, социальным развитием, религиозным развитием,
этиологическим развитием, этнологическим развитием, и кто знает чем еще. И потом беженцы от
Европейского геноцида и кочевые старожилы из Азии, с их различными генетическими маркерами Y-ДНК
хромосомы, сталкиваются друг с другом на краю Восточной Европы, и в течение 500 лет становятся
неотличимыми друг от друга. Одна культура, одна технология, единое общество, одно искусство,
керамика, религия, этиология, этнология, и все остальное. Как будто бы и не было Вавилонской башни,
которая разделила людей на враждующие фракции, как возвращение Эдема на Землю. А 500 лет спустя,
происходит еще одно чудо, волшебное разделения неотличимо одиноких людей на два контрастных и
противоположных множества, одно явное продолжение предыдущего единой кочевой животноводческой
археологической цивилизации, и другие чисто сельскохозяйственный, с языком несовместимой семьи, и
явно не связанное с предыдущей единой кочевой животноводческой археологической цивилизацией.
Генетически, преждевременно сказать, кто есть кто, такая задача является достаточно новой для
археологов. В остальном, и в том числе биологически, совершенно ясно, кто есть кто, и никакая масса
современных фальшивых ярлыков мне может изменить прошлую ситуацию в или на земле. |
Карта показывает, что нынешние носителями R1b в Центральной России распространилась до
Архангельска на Белом море. Очень вероятно, что это относительно недавний перемещение, хотя это еще
предстоит определить. В настоящее время среди жителей Европейской части России только около 5%
носители R1b.
Как было описано выше в этом исследовании, Арбины отправились на юг через Кавказ в Месопотамию и
на Ближний Восток около 6000 - 5000 лн, они создали Шумерскую цивилизацию, прошли на запад через
Египет до Атлантики, и через Гибралтарский пролив в Пиренеи. На их пути некоторые носители R1b-V88
откололись, углубились в Африку, и в настоящее время в заметных количествах населяют Камерун и Чад
(см. условные обозначения Рисунка 10, и описание и ссылки выше). К 4800 лн Арбины достигли
Иберии и стали первыми Колоколовидными Кубками. Эта дата была получена для общего предка субкладов
гаплотипов P312 и U106 (см. следующий раздел), и поддерживается данными археологии (Cardoso &
Soares, 1990; Martinez et al., 1996; Cardoso, 2001; Muller & Willigen, 2001; Nocete, 2006).
Несколько входных маршрутов гаплотипов R1b1a2 в Европе: из Восточно-Европейской
равнины, из Малой Азии/Ближнего Востока, из Пиренеев
Подводя итоги предыдущего раздела, Арбины входили в Европу с востока по нескольким маршрутам: из
Восточно-Европейской равнины (между Понтийскими степями и Балтийским морем); из Малой Азии и
Ближнего Востока, и после долгого пути вокруг Средиземноморского моря в Иберию, и дальше на север,
на Европейский континент. Первые два основных маршрутов были связаны с носителями субклада R-L23.
Иберийский маршрут был проделан главным образом людьми M269, из которых во время вступления на
Пиренеи выделились R-L51, и сразу же потом R-L11. Оба они очень похожи и время их общих
предков очень близко, как это показано в следующем разделе. 4850 лн из L11 сразу выделились два
“братских” субклада P312 и U106 (Клесов, 2011b), которые после долгой “популяционной горловины” на
грани исчезновения в конце концов выжили и расширились около 4000 - 3700 лн, а также активно
населили Европу, сначала как Колоколовидные Кубкиы, между 4000 и 3000 лн, а затем к эпохе Древнего
Рима, как Галлы и Кельты, упоминая только имена с присутствующими определенными историческими
“вехами”. На самом деле, в Европе жили десятки, если не сотни древних племен R1b.
98
Главные субклады R1b1a2 на Европейском континенте: Вход в Европу 4800 - 4500 лн
с востока и юго-запада
Почти все гаплотипы субклада R-M269 с датой 6000 - 5000 лн принадлежат либо к парагруппе R-M269*,
или ее нижестоящему субкладу R-L23. Арбины, которые мигрировали с востока в Европу в те времена, в
основном несли DYS393 = 12. В настоящее время мы видим в Европе только такие гаплотипы R1b1a2
(xL23), они происходят от общих предков живших максимум ~4500 лн. Либо потомки более древних предков
L23 не сохранились до настоящего времени, либо вовсе не пришли с Восточно-Европейской равнины, Малой
Азии и с Ближнего Востока.
Вышеприведенная схема показывает, что непосредственными нижестоящими субкладами Р-L23 были L51 и
L11 (ISOGG-2012, в сокращенном виде). Динамика этих субкладов гораздо лучше понимается через Иберию
в глубь континента, где миграция Арбинов в настоящее время отождествляется с Колоколовидными
Кубками.
Вопрос - где эти субклады L51 и L11 могли возникнуть? Если их “возраст” 6000-5000 лерт, они могли
бы отделиться в Малой Азии, на Ближнем Востоке или на Восточно-Европейской равнине, и оттуда попасть
в Европу. “Внутри-кладовые” гаплотипы, т.е. только субклад L51 или только L11, возможно, отражают
популяционные горловины, следовательно, они выглядят “моложе” своего реального возраста (в плане
мутаций и соответствующих TMRCA). Однако их “Внутри-кладовое” сравнение может выявить потеряное
(из-за популяционных горловин) время до более древних общих предков. Для анализа этих
субкладов, на Рисунке 11 показано комбинированное древо гаплотипов L51-L11.
99
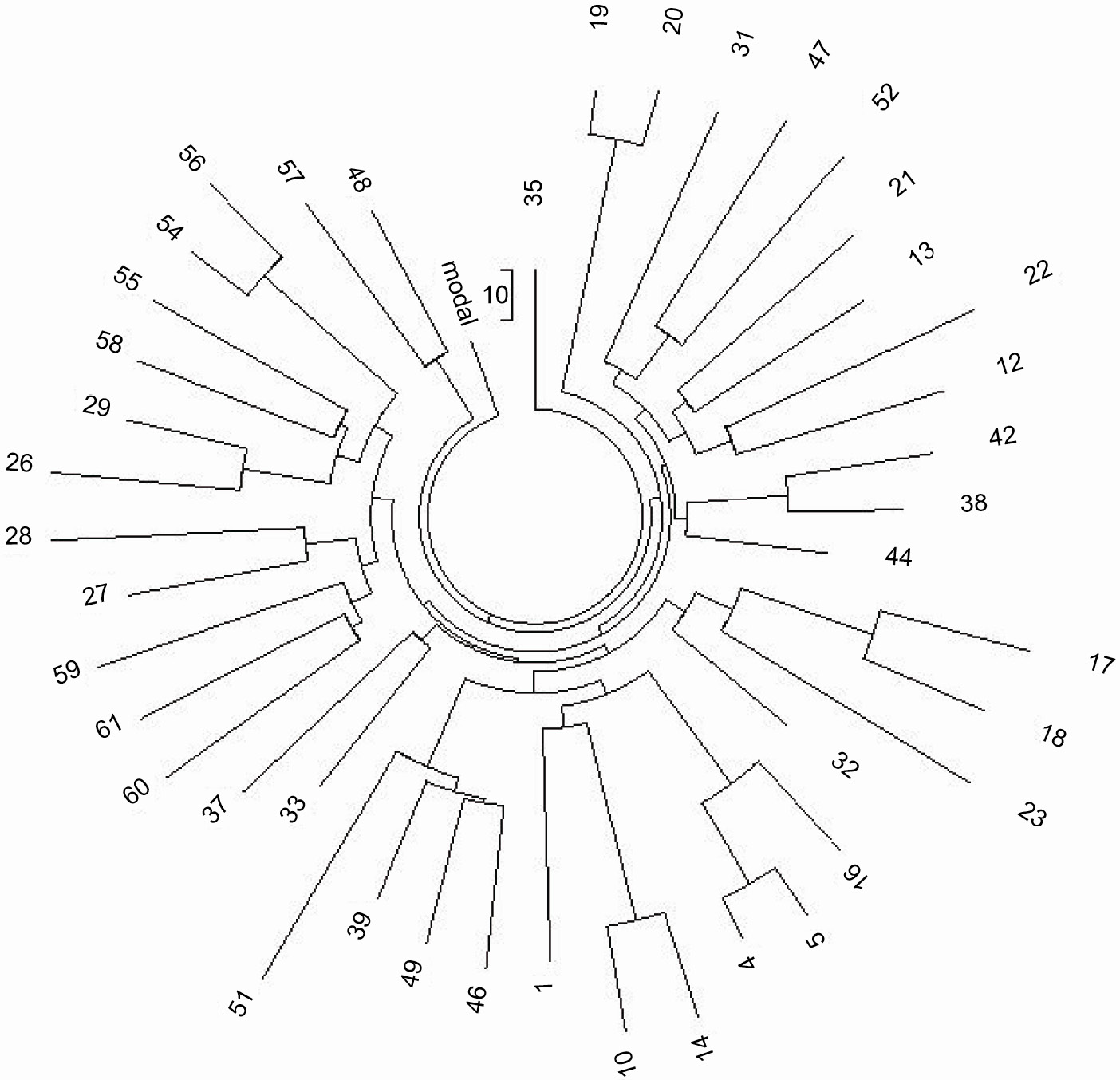
Одна треть гаплотипов на древе относятся к субкладу L51, и они занимают правую сторону. Другой
две трети гаплотипы L11 на левой стороне, и также дают некоторые “вставки” на ветви справа.
67-маркерные гаплотипы субкладов L51 и L11 очень похожи, поэтому в ряде случаев древо не смогло их
различить; пюэтому смешанные ветви. Древо также показывает, что “возрасты” двух субкладов также
очень похожи, так как “высота” (которая обычно указывает на “возраст") ветвей вокруг древа примерно
такая же. Базовый гаплотип субклада R-L51 выглядит следующим образом
13 24 14 11 11 14 13 12 12 13 13 29 - 17 9 10 11 11 25 15 19 30 15 16
17 18
- 11 11 19 23 16 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16
10 12 12 16 8 12 22 20 14 12 11 13 11 11 12 12 (L51)
Он отличается от базового гаплотипа R-L23 (5400±800 лн) на 9 мутаций (выше отмечены 12 мутаций,
но многие из них являются дробными), что дает 9/.12 = 75 → 81 поколений, или ~2025 лет “бокового”
расстояния. Все 15 гаплотипов содержат 280 мутаций от базового гаплотипа, что дает 280/15/.12 = 156
→ 184 поколений, или 4600±535 лет от их общего предка L51. В свою очередь, это дает (2025 + 5400 +
4600)/2 = 6000 лет, т.е. “возраст” общего предка обоих субкладов L23 и L51. Очевидно, это общий
предок самого субклада L23.
Иными словами, “возраст” субклада L51 4600±535 лет получен из мутаций ветви L51 на древе (Рис.
11) и подтвержден 1) мутационным расстоянием от базового гаплотипа родительского субклада
L23, 2) “возрастом” субклада L23 определенным ранее, и 3) общей филогениией субклада R1b. “Возраст”
субклада L51 хорошо вписывается во время прибытия Арбинов в Иберию (4800 лн), и в характер
распределения современных носителей L51 в Европе (самая высокая частота на Пиренеях, и затем сразу
же на севере Франции и на островах, см. [Myres et al, 2010]. Субклад L51 практически отсутствует в
Юго-Восточной, Восточной и Северо-Восточной Европе (там же).
Анализ субклада L11 более сложной, поскольку его гаплотипы разбросаны по всему древу, по крайней
мере четырьмя ветвями, каждая со своим общим предком. Они отличаются друг от друга на 32 мутаций,
которые дают 32/4/.12 = 67 → 72 поколений, или 1800 лет ниже их среднего “возраста” ~2500 лет. Таким
образом, “возраст” субклада R-L11 составляет ~4300 лет, что действительно соответствует 4600±535 лет
для субклада L51. Базовый гаплотип субклада L11 выглядит следующим образом:
13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 30 15 15 17 17 -
11 11 19 23 16 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12
22 20
13 12 11 13 11 11 12 12 (L11)
Пять отклонений между ними (выделены жирным шрифтом) фактически дают 3,7 мутаций (поскольку
большинство из них являются дробными), что дает 3.7/.12 = 31 → 32 поколений, то есть всего 800
“боковых” лет между ними. Это подтверждает, что субклады L51 и L11 очень близки друг к другу по
времени. Их общий предок (который предположительно должны быть L51) жил (800 + 4600 + 4300)/2 = 4850
лн. Кажется, что L11 откололся только ~250 лет спустя. Скорее всего, они все создали археологическую
культуру Колоколовидных Кубков. Самый древние артефакты Колоколовидных Кубков были найдены в
Португалии, даторованные от 4800 - 4600 лн (Cardoso & Soares, 1990; Martinez et al., 1996; Cardoso,
2001; Muller & Willigen, 2001; Nocete, 2006).
100
Главные субклады R1b1a2 на Европейском континенте: Население Европы после 4800
лн, главные субклады на островах и их вероятное происхождение
Филогения R1b (см. диаграмму выше) показывает, что нижестоящими субкладами Р-L11 являются
субклады P312 и U106. Их 67-маркерные гаплотипы следующие:
13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 - 11 11 19 23 15 15 18
17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12
12 (P312)
13 23 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 - 11 11 19 23
16 15 17 17 37 39 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12
22 20 13 12 11 13 11 11 13 12 (U106)
Между ними насчитывается 5,5 мутаций (выделены жирным шрифтом) (DYS456 в P312 колеблются между 15
и 16 в различных наборах данных), что отделяет два базовых гаплотипа на 5.5/.12 = 46 → 48 поколений,
или 1200 лет. Поскольку общие предки обоих субкладов жили 4350±700 и 4175±430 лн (Клесов, 2011b), их
общий предок жил (1200 + 4350 + 4175)/2 = 4850±700 лн. Действительно, это “возраст” субклада L11 в
пределах погрешности. В общем, безопасно принять “возраст” обоих P312 и U106 как ~4200 лн, так как
оценки для разных наборов данных колеблются вокруг этого значения. Между ними существует 1200 лет
разницы (см. выше), что определяет время до их общего предка L11 как (4200 + 4200 + 1200)/2 = 4800
лн.
Как уже было сказано выше, четыре субклада L51, L11, P312 и U106 исторически представляют собой
первую волну движения Колоколовидных Кубков из Иберии на Европейский континент. Наиболее высокая
частота субклада P312* в Европе в настоящее время наблюдается в Иберии (Myres et al, 2010), однако,
наивно судить о древних местах субкладов на основе их сегодняшнего распределения
(Хотя это почему-то рутинно применяется к динамической ситуации в Восточной Европе). Их
перемешивание на континенте в последние тысячелетия было слишком интенсивным для однозначной
экстраполяции. В настоящее время региональное распределение нижестоящих субкладов R-M269,
мигрировавших с Пиренеев почти пять тысячелетий назад, является показательно неинформативным, за
исключением случаев некоторых местных субкладов. Один из них - это R-M222.
Похоже, что история появления R-M222 на островах возможно началась с приплытия носителей L11* из
Иберии на острова около 4500 - 4000 лн. Однако, это всего лишь одна из теорий. Другая основана на
миграции носителей L21 на острова по суше. И тогда P312, нижестоящий клад от L11, также интенсивно
представленный на островах вместе с его нижестоящими субкладами, а также его "братский” субклад U106
и нижестоящие клады (там же). L21, нижестоящий клад от P312, изобилен на островах, в отличие от
своего “братского” субклада U152 (там же). Эта древняя миграция Арбинов из Иберии на острова
объясняет некоторые данные о генетике “Испанского происхождения Ирландцев”.
M222, нижестоящий клад от L21, является одним из наиболее представленных субкладов R1b на
островах. Массивный Проект FTDNA Ирландское Наследие (см. Приложение) перечисляет несколько
тысяч гаплотипов R1b (с субкладами) на островах, из которых M222 является крупнейшим и включает в
себя около 25% всех R1b (Клесов, 2010е). Субклад наиболее выражен в Ирландии. В Шотландии он
наблюдается в основном в низинах и в центральной области, и в Англии предствлен плохо. Базовый
гаплотип субклада M222:
13 25 14 11 11 13 12 12 12 13 14 29 - 17 9 10 11 11 25 15 18 30 15 16 16 17 - 11 11 19 23 17 16 18
17 38 39 12 12 - 11 9 15 16 8 10 10 8 10 10 12 21 23 16 10 12 12 16 8 12 25 20 13 12 11 13 11 11 12
12 (M222)
Интересно, что базовый гаплотип выше отличается от “братских” базовых гаплотипов P312 и U106 на
18 мутаций в каждом случае (на самом деле 15 мутаций, так как некоторые мутации дробные). Это
показывает, что P312 и U106 действительно очень близкие (во времени и в происхождении) базовые
гаплотипы, и что их общий предок, носитель базового гаплотипа L11, по времени жил очень близко к ним
обоим. 15 мутаций относят их общего предка на 4600±500 лн.
Многочисленные анализы данных гаплотипов M222 производят временный промежуток сегдняшних потомков
M222 до их общего предка как 1450±160 лет (Клесов, 2010П). Тем не менее, перекрестное сравнение
многочисленных линий субклада M222 показало, что линия M222 возникла не позднее, чем в начале нашей
эры, около 400 лет раннее чем оценка выше. Это указывает на возможную популяционную горловину между
временем появления M222 и расширения субклада в середине 1-го тысячелетия нашей эры. Нельзя
определить, по крайней мере пока, где конкретно субклад M222 появился впервые (то есть единичная
нуклеотидная мутация G → А произошла в определенном нуклеотиде Y-хромосомы определенного
человека, носителя вышестоящего субклада L21), будь то на островах, в Северо-Западной Европе, или в
другом месте. Мы также не можем определить, с чем конкретно прямые потомки R-M222 боролись за их
выживание на протяжении нескольких столетий. Были ли это непрерывные климатические события? Римские
вторжения? Эпидемии? Голод? или их комбинация? Мы знаем, однако, что в конце концов это были
острова, где мутация в Y-хромосоме выжила и процветала.
Филогенетическая диаграмма показывает, что мутация M222 возникла одновременно с его “братскими”
SNP-мутациями, которые определяют теперь субклады L226 и P314. Первый из них имеет общего предка
жившего 1500±170 лн, т.е. в то же время что и предок M222, хотя мутация в субкладе L226 была C → T.
Его базовый гаплотип выглядит следующим образом:
13 24 14 11 11 14 12 12 11 13 13 29 - 17 8 9
11 11 25 15 19 29 13 13 15 17 - 11 11 19 23 15 15 18 17
36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 15 10 12 12 15 8 12 22 20 13
12 11 13 11 11 12 12 (L226)
Он сильно отличается 21-й мутацией от своего “братского” субклада M222, несмотря на данные что их
общие предки жили в одно и тоже время. Это только подтверждает что их общий предок, основатель
субклада L21, жил около 4100 лн. Это также показывает, что L21 отпочковался от P312 практически
сразу (см. филогенетическую диаграмму), так как P312 возник 4350 - 4200 лн, и L21 около 4100 лн. Их
67-маркерные гаплотипы одинаковы, в небольшими дробными мутационными различиями:
13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 - 11 11 19 23 15 15 18
17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12
12 (P312)
13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 - 11 11 19 23
16 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11
13 11 11 12 12 (L21)
101
Единственное кажущееся различие, в DYS456, также является дробным, так как в P312 этот аллель
колеблется между значениями 15 и 16. Несмотря на подобные хронологии, за предыдущие 2600 лет
субклады M222 и L226 имели совершенно разные истории. А еще один “братский” субклад, P314 (точнее,
P314.2, так как та же SNP-мутация T → C P314 произошла в гаплогруппе H2a), несколько “старше”, с его
2225±300 лн, однако он может уложиться в те же сроки когда действительно возник M222. Его базовый
гаплотип
13 23 14 11 11 14 12 13 13 13 13 29 - 17 9 10 11 11 26 15 19 29 15 15 16 17 - 10 11 19 23 16 15 18
18 37 38 12 12 - 11 9 16 16 8 10 10 8 11 10 12 23 23 17 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12
12 (P314.2)
снова выделяется по сравнению с базовыми гаплотипами субкладов M222 и L226 на 24 и 17 мутаций
соответственно. В семье L21 это третья, совершенно отдельная линия. 10 полных мутаций от
родительского базового гаплотипа L21 производят 10/.12 = 83 → 91 поколений, или 2275 лет
мутационного различия, и помещают начало субклада L21 на (2275 + 4100 + 2225)/2 = 4300 лет, что
находится в пределах погрешности 4100 лет просчитанных выше. Все эти базовые гаплотипы, вместе с
внутри-кладными и меж-кладными датировками, образуют довольно здравую систему ДНК-генеалогии
Арбинов в Европе и Азии.
Материалы и Методы
Четыре тысячи четыреста восемь (4408) гаплотипов R1b (с субкладами) были собраны в базового
данных из FTDNA, YSearch, и в экспертно рецензированых научных публикациях.
Методология анализа гаплотипов из наборов данных была описана в предыдущей публикации в этом
журнале [Рожанский и Клесов (Rozhanskii & Klyosov), 2011]. В этом исследовании используются линейные
и логарифмические методы; последний когда базовый гаплотип в наборе данных был легко
идентифицирован, как описано в (Клесов, 2009c). Константы скорости мутаций перечислены в (Клесов,
2009с; Рожанский и Клесов, 2011), и в ряде случаев приведены в тексте этого документа. Наиболее
широко используемыми константами скорости мутаций, в том числе используемые в данной работе,
являются следующие (количество мутаций на гаплотип на условное поколение в 25 лет); ссылки указывают
примеры гаплотипов в указаном формате:
0,013, 8-маркерный гаплотип (Zhong et al.).
0,017, 8-маркерный (Contu et al.).
0,018, 10-маркерный (Myres et al.).
0,020, 11-маркерный (Cruciani et al.).
0,019, 11-маркерный (Zalloua et al.).
0,020, 12-маркерный в формате FTDNA
0,024, 12-маркерный (Малярчук и др.).
0,034, 17-маркерный в формате Y-Filer (Herrera)
0,035, 19-маркерный (Балановский и др.).
0,046, 25-маркерный в формате FTDNA
0,090, 37-маркерный в формате FTDNA
0,120, 67-маркерный в формате FTDNA
0,198, 111-маркерный в формате FTDNA
Форматы гаплотипов FTDNA приведены в
http://www.familytreedna.com/faq/answers.aspx?id=9
Древа гаплотипов были составлены с использованием программы PHYLIP, Программный Пакет Выделения
Филогении (см. Клесов, 2009c, 2009d и ссылки там). Коррекция на обратные мутацийи были введены как
описано в (Клесов, 2009c). Погрешности были рассчитаны, как описано в (Клесов, 2009c).
Базовые гаплотипы в наборе данных были определены путем минимизации мутаций; по определению,
базовый гаплотип имеет минимальные коллективное число мутаций в наборе данных, происходящих от
одного общего предка. Базовый гаплотип - это предковый гаплотип или близкое приближение к нему.
Промежуток времени до общего предка двух базовых гаплотипов определяется следующим образом:
1) подсчитывается количество мутаций между двумя базовыми гаплотипами,
2) полученное число делится на константу скорости мутаций,
3) вводится поправка на обратные мутации, рассчитываемые по следующей формуле (Клесов, 2009c):
λ = λobs/2obs(1+exp2(λobs)) (1)
где:
λobs = наблюдаемое среднее число мутаций на маркер в наборе данных (или в ветви, если набор данных
содержит несколько ветвей/линий),
λ = среднее (фактическое) число мутаций на маркер с поправкой на обратные мутации,
4) добавляется полученное значение, умноженное на 25 (лет), что представляют собой “боковой”
промежуток времени между моментами появления двух базовых гаплотипов, к TMRCA
(Time to Most Recent Common Ancestor/Время до Самого Недавнего Общего
Предка = ВСНОП) базового гаплотипа и делим на 2.
Результатом является TMRCA (время до самого недавнего общего предка) для рассмотренных двух базовых
гаплотипов. Следующие примеры основаны на конкретных случаях в данном исследовании.
Пример 1: Расчет отрезка времени до общего предка ветви (самой удаленной на древе гаплотипов
показаном на
Рисунке 1), который содержит 12 из 8-маркерных гаплотипов с коллективными 65 мутациями от их
базового гаплотипа. Поскольку константа скорости мутаций этих гаплотипов равна 0.013 на гаплотип за
условное поколение (25 лет), мы имеем 65/12/.013 = 417 → 619 поколений, то есть 619 × 25 = 15.475
лет до общего предка ветви. Стрелка показывает коррекцию на обратные мутации. Эта поправка может
быть рассчитана по формуле (1) следующим образом. Так как наблюдаемое число мутаций на маркер
65/12/8 = 0,677, мы используем формулу (1) и получаем
1/2(1+exp(0.677))λobs
Полученное число 1,484 является поправочным коэффициентом для обратной мутации. Таким образом,
путем умножения (нескорректированного числа поколений) 417 ×
1,484, мы получаем скорректированное число поколений 619, то есть 619 × 25 = 15.475 лет. Это обычно
изображается 417 → 618 (поколений). Так как на 65 мутаций в наборе данных погрешность составляет
15,93% (рассчитано как описано в Клесов, 2009c), мы наконец получим отрезок времени до общего предка
гаплотипов равный 15475±2500 лет.
102
Пример 2: Два 67-маркерных базовых гаплотипа субкладов R-P312 и R-U106 отличаются на 5,5 мутаций
(см. текст выше). Применяя правила объясненные в предыдущем примере, мы получаем 5.5/.12 = 46 → 48
поколений, то есть 1200 лет от общего предка двух базовых гаплотипов (то есть, двух субкладов). В
случае, если бы два базовых гаплотипа имели одинаковый “возраст” (то же самое TMRCA), их общий
предок жил бы 600 лет “глубже” во времени от любого из них. Но в данном случае два TMRCA равны 4350
и 4175 годам соответственно. Поэтому их общий предок жил (1200 + 4350 + 4175)/2 = 4850 лет назад.
Заключение
Результаты этого исследования поддерживают теорию что гаплогруппа R1b возникла в Центральной
Азии, по-видимому, в Южной Сибири и соседних областях, около 16.000 лет назад. Предыдущая история
гаплогруппы напрямую связана с появлением Европеоидов (Кавказоидов) ~58000 лн, скорее всего, в
огромном треугольнике простирающемся от Западной Европы через Восточно-Европейскую равнину на восток
и в Левант на юг, как это было предложено в (Клесов, 2011d). Последующая последовательность
SNP-мутаций в Y-хромосоме в ходе их миграции на восток к Южной Сибири, с появлением гаплогрупп
NOP ~48000 лн и Р ~38000 лн привела в конечном итоге к гаплогруппе R ~ 30000 лн и R1 ~26000 лн, а
затем к гаплогруппам R1a/R1a1 ~20,000 лн (время между появлением R1a и R1a1 неопределенно) и R1b
~16000 лн (там же).
На основании фонетических соответствий, мы называем носителей R1a Ариями
(от Aran, тот же формат как и Арбин), и носителей R1b Арбинами. В первом случае это
название оправдано, так как носители гаплогруппы R1a стали легендарными Ариями, прибывшими в
Индостан и на Иранское нагорье ~3500 лн. Другими словами, эти Арийцы принадлежали к гаплогруппе R1a,
следовательно, двойной смысл (хотя и совпадающий) термина Арийцы. Арбины - это удобный общий термин,
избегающий повторения: “носители гаплогруппы R1b”.
В какой-то момент, Арбины начали миграцию на запад, через Среднюю Азию, Северный Казахстан, Южный
Урал, к Восточно-Европейской равнине, где между 12.000 и 4500 лн они создали ряд археологических
культур (видимо, в том числе Сероглазовскую, Хвалынскую, Самарскую, Средневолжскую, Срубную,
Катакомбную, а также “Прото-Куранную” и/или “Курганную” культуры, последняя в значительной степени
оспаривается и не принимается многими историками; следует подчеркнуть, что все эти приведенные выше
соображения в отношении археологических культур в настоящее время можно рассматривать только как
очень предварительные сопоставления) (причина предварительности не в
отсутствии генетически связывающих эти культуры археологических материалов, а полное отсутствие
палеогенетических исследований этих археологических культур). “Арбины” мигрировали на юг,
около 6000 лн часть мигрировала через Кавказ в Анатолию (оставляя позади их гаплогруппы R1b и
соответствующие гаплотипы); в остальную Малую Азию и на Ближний Восток. Видимо, Арбины создали
Шумерскую культуру и государство, и по нескольким маршрутам мигрировали на запад в Европу, в
основном неся субклад R-M269 и его нижестоящий субклад L23. Один маршрут был по северной дуге, с
Восточно-Европейской равнины на запад ~ 4600 - 4400 лн, другой одновременный маршрут с теми же двумя
субкладами шел на запад вдоль Малой Азии и Ближнего Востока, и еще один маршрут, наибольше
заполнивший Европу, шел вдоль Северной Африки-Средиземного моря через Древний Египет до Пиренеев,
прибыв ~4800 лн. На этом маршруте, племя R1b-V88 разделилось и часть пошла на юг, в конечном итоге в
Центральную Африку (в основном, Камерун и Чад, судя по их современному распределению), где общий
предок современноо R1b-V88 гаплотипов жил ~4400 лн.
Во время прибытия к Иберии ~4800 лн, субклад M269 отпочковал клад M51, и вскоре после этого
отпочковал клад L11 и нижестоящие субклады. Они стали племенами Колоколовидных Кубков и двинулись на
север вместе с вновь возникшими субкладами P312 и L21, последний возник несколько веков после P312.
Те субклады и их нижестоящие клады эффективно и без больших разрывов населили Европу от Атлантики до
Балкан, Карпат, сегодняшней Польши, западной границы Восточно-Европейской равнины, до Балтийского
моря, о чем свидетельствуют гладкие древа гаплотипов показывающие беспрерывное распространение
гаплотипов R1b.
На Островах история гаплотипов R1b и линий субкладов была другой. Носители L11, L21 и P312
мигрировали на острова по суше и по морю одновременно с теми Арбинами, которые заполняли Европу
между 4000 и 2500 лн, и сформировали соответствующие “местные” субклады, такие как P314, M222, L226,
которые в основном заселили Острова. В результате, значительная часть Островов заселена почти
исключительно Арбинами, с частотой их аллелей достигающей 92% - 96% населения. В целом, пропорция
Арбинов в Западной и Центральной Европе достигает, хотя и не равномерно, около 60% населения.
Это исследование по существу представляет собой пример применения ДНК-генеалогии для изучения
истории человечества. Этот пример — сложная и требовательная задача, которая также затрагивает
некоторые таинственные загадки истории и лингвистики. Одна из этих головоломок то, что язык или
языки, на которых говорили Арбины от 16000 до 3000 лн, и это почти наверняка был долговременный в
его динамике не-Индо-Европейский язык. В мнении лингвистов принято, что эти языки разрозненные и
бессвязные “мертвые” и не совсем мертвые языки, такие как прото-Тюркский, Шумерский,
Северо-Кавказский, Дене-Кавказской, Баскский, и многие до-Индоевропейские языки Европы периода 5000
- 2000 лн, а также некоторые более поздние языки (M.Gimbutas назвала эти
языки “Старой Европой”, и приписала их оседлым земледельческим населениям; к сожалению, она не
уловила, что “Старая Европа” - это смесь предыдущих охотников-собирателей и земледельческого
населения“Старий Европы” с Азиатскими мигрантами охотников-собирателей отмеченных маркером R1a1,
которые достигли Балкан ок. 10-8 тыс. до н.э., и наряду с другими инновациями принесли их
до-Курганную этиологию с богиней Пра-мамой). Возможно, язык Арбинов был первоначально
единым языком легко протекающим через тысячелетия и по всей Евразии. Впрочем, это тема другого
исследования.
Благодарности
Автор выражает благодарность Susan Hedeen за ее ценную помощь в подготовке рукописи.
Библиография
Balanovsky, O., Dibirova, K., Dybo, A., Mudrak, O., Frolova, S., Pocheshkhova, E.
et al. (2012). Parallel evolution of genes and languages in the Caucasus region. Molecular
Biology and Evolution, 29, 359-365.
Balaresque, P., Bowden, G. R., Adams, S. M., Leung, H.-Y., King, T. E., Rosser, Z. H. et al. (2010).
A predominantly Neolithic origin for European paternal lineages. PLoS Biology, 8, 1000285
Barac, L., Pericic, M., Klaric, I. M., Janicijevic, B., Parik, J., Rootsi, S. et al. (2003a). Y
chromosome STRs in Croatians. Forensic Science International, 138,
127-133.
Barac, L., Pericic, M., Klaric, I. M., Rootsi, S., Janicijevic, B., Kivisild, T. et al. (2003b). Y
chromosomal heritage of Croatian population and its island isolates. European Journal of Human
Genetics, 11,
535- 542. doi:10.1038/sj.ejhg.5200992
Bengtson, J. D. (1997). The riddle of Sumerian: A Dene-Caucasian language? Mother Tongue, 3,
63-74.
Capelli, C., Brisighelli, F., Scarnicci, F., Arredi, B., Caglia, A., Vetrugno, G. et al. (2007). Y
chromosome genetic variation in the Italian peninsula is clinal and supports an admixture model for
the Mesolithic-Neolithic encounter. Molecular Phylogenetics and Evolution, 44, 228-239.
doi:10.1016/j.ympev.2006.11.030
Cardoso, J. L. (2001). Le phenomene campaniformedans les basses vallees du Tage et du Sado
(Portugal). In F. Nicolis (Ed.), Bell Beak-ers Today, Trento: Ufficio Beni Archeologici.
103
Cardoso, J. L., & Soares, A. M. (1990). Chronologia absoluta para o campaniforme da Estremadura e
do Sudoeste de Portugal. O Arqueologo Portugues, 8-10, 203-228.
Cinnioglu, C., King, R., Kivisild, T., Kalfoglu, E., Atasoy, S., Cavalleri, G. L. et al. (2004).
Excavating Y-chromosome haplotype strata In Anatolia. Human Genetics, 114, 127-148.
doi:10.1007/s00439-003-1031-4
Contu, D., Morelli, L., Santoni, F., Foster, J. W., Francalacci, P., Cucca, F. (2008). Y-Chromosome
based evidence for pre-Neolithic origin of the genetically homogeneous but diverse Sardinian
population: inference for association scans. PLoS ONE, 3.
doi:10.1371/journal.pone
Cruciani, F., Trombetta, B., Sellitto, D., Massaia, A., Destro-Bisol, G., Watson, E. et al. (2010).
Human Y chromosome haplogroup R-V88: a paternal genetic record of early mid Holocene trans-Saharan
connections and the spread of Chadic languages. European Journal of Human Genetics, 18,
800-807. doi:10.1038/eihg.2009.231
Di Gaetano, C., Cerutti, N., Crobu, F., Robino, C., Inturri, S., Gino, S. et al. (2009).
Differential Greek and northern African migrations to Sicily are supported by genetic evidence from
the Y chromosome. European Journal of Human Genetics, 17, 91-99.
doi:10.1038/ejhg.2008.120
Hammer, M. F., Behar, D. M., Karafet, T. M., Mendez, F. L., Hallmark, B., Erez, T. et al. (2009).
Extended Y chromosome haplotypes re-solve multiple and unique lineages of the Jewish priesthood.
Human Genetics, 126, 707-717.
doi:10.1007/s00439-009-0727-5
Herrera, K. J., Lowery, R. K., Hadden, L., Calderon, S., Chiou, C., Yepiskoposyan, L. et al. (2011).
Neolithic patrineal signals indicate that the Armenian plateau was repopulated by agriculturalists.
European Journal of Human Genetics, 20, 313-320.
doi:10.1038/ejhg.2011.192
Kang, L., Lu, Y., Wang, C., Hu, K., Chen, F., Liu, K. et al. (2011). Y-chromosome O3 haplogroup
diversity in Sino-Tibetan populations reveals two migration routes into the Eastern Himalayas.
Annals of Human Genetics, 76, 92-99.
doi:10.1111/j.1469-1809.2011.00690
Klyosov, A. A. (2008a). Mysteries of the “Western European” haplogroup R1b.
Proceedings of the Russian Academy of DNA Genealogy, 1, 568-630 (in Russian).
Klyosov, A. A. (2008b). Origin of the Jews via DNA genealogy. Proceedings of the Russian Academy
of DNA Genealogy, 1, 54-232.
Klyosov, A. A. (2009a). A comment on the paper: Extended Y chromosome haplotypes resolve multiple
and unique lineages of the Jewish priesthood. Human Genetics, 126, 719-724.
doi:10.1007/s00439-009-0739-1
Klyosov, A. A. (2009b). Once again on the “population mutation rate” of L. Zhivotovsky, or how
fables are born. Proceedings of the Russian Academy of DNA Genealogy, 2, 1162-1181 (in
Russian).
Klyosov, A. A. (2009c). DNA Genealogy, mutation rates, and some historical evidences written in
Y-chromosome. I. Basic principles and the method. Journal of Genetic Genealogy, 5,
186-216.
Klyosov, A. A. (2009d). DNA Genealogy, mutation rates, and some historical evidences written in
Y-chromosome. II. Walking the map. Journal of Genetic Genealogy, 5,
217-256.
Klyosov, A. A. (2010a). Haplogroup R1b1 and its subclades in
Asia. Proceedings of the Russian Academy of DNA Genealogy, 3, 1676 - 1695 (in
Russian).
Klyosov, A. A. (2010b). Haplotypes of haplogroup R1b in central Africa. Proceedings of the
Russian Academy of DNA Genealogy, 3, 369-378 (in Russian).
Klyosov, A. A. (2010c). Haplogroup R1b. Part 2. Proceedings of the Russian Academy of DNA
Genealogy, 3, 406-475 (in Russian).
Klyosov, A. A. (2010d). The “age” of subclade R1b1a2-M269 and its subclades (L23, L51, L11).
Proceedings of the Russian Academy of DNA Genealogy, 3, 1310-1315 (in Russian).
Klyosov, A. A. (2010e). Irish haplotypes and haplogroups. Proceedings of the Russian Academy of
DNA Genealogy, 3, 1029-1053.
Klyosov, A. A. (2011a). Origin of ancient subclades of
haplogroup R1b — Regions and times. Proceedings of the Russian Academy of DNA Genealogy,
4, 2227-2245 (in Russian).
Klyosov, A. A. (2011b). Haplotypes of R1b1a2-P312 and related subclades: Origin and “ages” of most
recent common ancestors. Proceedings of the Russian Academy of DNA Genealogy, 4, 1127-1195.
Klyosov, A. A. (2011c). Haplogroups and haplotypes in Armenia (haplogroups J2, R1b-L23, R1b-M269,
and T-M184). Proceedings of the Russian Academy of DNA Genealogy, 4, 1985-1993 (in Russian).
Klyosov, A. A. (2011d). “Out of Africa” theory re-examined. Proceedings of the Russian Academy of
DNA Genealogy, 4, 1908-1977 (in Russian).
Klyosov, A. A., & Rozhanskii, I. L. (2012a). Haplogroup R1a as the Proto-Indo-Europeans and the
legendary Aryans as witnessed by the DNA of their current descendants.
Advances in Anthropology, 2,
1- 13. doi:10.4236/aa.2012.21001
Klyosov, A. A., & Rozhanskii, I. L. (2012b). Re-examining the “out of Africa” theory and the origin
of Europeoids (Caucasoids) in light of DNA genealogy. Advances in Anthropology, 2, in press.
Kramer, S. N. (1971). The Sumerians: Their history, culture, and character. Chicago, IL:
University of Chicago Press.
Lashgary, Z., Khodadadi, A., Singh, Y., Houshmand, S. M., Mahjoubi, F., Sharma, P. et al. (2011). Y
chromosome diversity among the Iranian religious groups: A reservoir of genetic variations.
Annals of Human Biology, 38, 364-372.
Lobov, A. S. (2009). Structure of the gene pool of the Bashkir subpopulations. Ufa.
http://ftp.anrb.ru/molgen/Lobov_AS.PDF
Mallory, J. P. (1989). In search of the indo-Europeans: Language, archaeology and myth.
London: Thames and Hudson.
Malyarchuk, B., Derenko, M., Denisova, G., Maksimov, A., Wozniak, M., Grzybowski, T. et al. (2011).
Ancient links between Siberians and native Americans revealed by sub-typing the Y chromosome
haplogroup Q1a. Journal of Human Genetics, 56, 583-588
Martinez, P. V., Lull, V., & Mico, R. (1996). El vaso campaniforme, in: Cronologia de la
Prehistoria Reciente de la Peninsula Iberica y Baleares (c. 2800-900 cal. A.N.E.). British
Archaeological Reports, International Series 652, 105-110.
Morelli, L., Contu, D., Santoni, F., Whalen, M., Francalacci, P., & Cucca, F. (2010). A comparison
of Y-chromosome variation in Sardinia and Anatolia is more consistent with cultural rather than
demic diffusion of agriculture. PLoS ONE, 5, e10419,
doi:10.1371/journal.pone.0010419
Muller, J., & Van Willigen, S. (2001). New radiocarbon evidence for European Bell Beakers and the
consequences for the diffusion of the Bell Beaker phenomenon. In F. Nicolis (Ed.),
Bell Beakers Today, Trento: Ufficio Beni Archeologici.
Myres, N. M., Rootsi, S., Lin, A. A., Jarve, M., King, R. J., Kutuev, I. et al. (2010). A major
Y-chromosome haplogroup R1b holocene era founder effect in central and western Europe. European
Journal of Human Genetics, 19, 95-101.
Nocete, F. (2006). The first specialized copper industry in the Iberian Peninsula: Cabezo Jure
(2900-2200 BC). Antiquity, 80, 646-657.
Pericic, M., Lauc, L. B., Klaric, A. M. et al. (2005). High-resolution phylogenetic analysis of
southeastern Europe traces major episodes of paternal gene flow among Slavic populations.
Molecular Biology and Evolution, 22, 1964-1975.
doi:10.1093/molbev/msi185
Roewer L., Willuweit S., Kruger C., Nagy M., Rychkov S., Morozowa I. et al. (2008). Analysis of Y
chromosome STR haplotypes in the European part of Russia reveals high diversities but
non-significant genetic distances between populations. International Journal of Legal Medicine,
122, 219-223. doi:10.1007/s00414-007-0222-2
Rozhanskii, I. L., & Klyosov, A. A. (2011). Mutation rate constants in DNA genealogy (Y chromosome).
Advances in Anthropology, 1, 26-34.
doi:10.4236/aa.2011.12005
Semino, O., Passarino, G., Oefner, P. J., Lin, A. A., Arbuzova, S., Beckman, L. E. et al. (2000).
The genetic legacy of paleolithic Homo sapiens sapiens in extant Europeans: A Y chromosome
perspective. Science, 290, 1155-1159.
doi:10.1126/science.290.5494.1155
Underhill, P. A., Myres, N. M., Rootsi, S., Metspalu, M., Zhivotovsky, M. A., King, R. J. et al.
(2009). Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup
R1a. European Journal of Human Genetics, 18,
479-484.
105
Weale, M. E., Yepiskoposyan, L., Jager, R. F., Hovhannisyan, N., Khudoyan, A.,
Barbage-Hall, O. et al. (2001). Armenian Y chromosome haplotypes reveal strong regional structure
within a single ethnonational group. Human Genetics, 109, 659-674.
doi:10.1007/s00439-001-0627-9
Wells, R. S., Yuldasheva, N., Ruzibakiev, R., Underhill, P. A., Evseeva, I., Blue-Smith, J. et al
(2001). The Eurasian heartland: a continental perspective on Y-chromosome diversity. Proceedings
of the National Academy of Sciences USA, 98, 10244-10249.
doi:10.1073/pnas.171305098
Wiik, K. (2008). Where did European men come from? Journal of Genetic Genealogy, 4,
35-85.
Zakharov, S. V. (2010). On the origin of Ботайскую culture. Vestnik of Archaeology, Anthropology,
and Ethnography, 1, 49-58.
Zalloua, P. A., Xue, Y., Khalife, J., Makhoul, N., Debiane, L., Platt, D. E. et al. (2008).
Y-Chromosomal diversity in Lebanon is structured by recent historical events. American Journal of
Human Genetics, 82,
973-882.
doi:10.1016/j.ajhg.2008.01.020
Zhivotovsky, L. A., Underhill, P. A., Cinnioglu, C., Kayser, M., Morar, B., Kivisild, T. et al.
(2004). The effective mutation rate at Y-chromosome short tandem repeats, with application to human
population divergence time. American Journal of Human Genetics, 74,
50-61. doi:10.1086/380911
Zhong, H., Shi, H., Qi, X.-B., Duan, Z.-Y., Tan, P.-P., Jin, L. et al. (2010). Extended Y-chromosome
investigation suggests post-Glacial migrations of modern humans into East Asia via the northern
route. Molecular Biology and Evolution, 28, 717-727.
APPENDIX
The following DNA projects were selected as primary haplotype databases:
http://www.familytreedna.com/public/R1b1b1/default.aspx?section=yresults
http://www.familytreedna.com/public/ht35new/default.aspx
http://www.familytreedna.com/public/AssyrianHeritageDNAProject/default.aspx?section=yresults
http://www.familytreedna.com/public/IrelandHeritage/default.aspx?section=yresults
http://www.familytreedna.com/public/R-L21/default.aspx?section=yresults
Reference data were selected according to SNP assignment from YSearch database: (http://www.ysearch.org)
and public projects of FTDNA (http://www.familytreedna.com)
|