| 101 Summary
This article attempts to merge the data of contemporary linguistics and DNA
genealogy in order to describe the migrations and settlement of peoples and languages in Europe
after the last Ice Age. In the new paradigm, three important groups of players have been identified:
— R1a haplogroup bearers, conditionally identified as Aryans. They arose around 20,000 years before
the present (ybp) in central Asia and the Altai Mountains; after their migration along the southern
route, they arrived in Europe between 10,000 - 9000 ybp, bringing proto-Indo European (PIE) and Indo
European (IE) languages. In 4800 ybp they migrated eastward from Europe to the Russian Plane and
then to India. About 3000 - 2500 ybp they migrated with their IE languages from the Russian Plain
back to central, western, and southern Europe, laying the genetic groundwork for peoples later
called Celts, Germans, Italics, Greeks, Illyrians, and Balto-Slavs. — E, F, G, J, I, K haplogroup
bearers. The dates of their arrival in Europe (sometime before 5000 ybp) and their migration routes
remain obscure. They apparently spoke non-IE languages. — R1b haplogroup bearers, called the Arbins.
They arose about 16,000 ybp in central Asia, and migrated to Europe along a northern route. They
arrived in Europe between 4800 and 4500 ybp bringing with them several non-IE languages. It seems
that the arrival of the Aryans (R1a) in Europe was peaceful. There are no clear indications that
their arrival triggered any sort of violence. However, the migration of the Arbins (R1b) was marked
by an almost complete elimination of the E1b, F, G2a, J, I1, I2, and K haplogroups from Europe. Our
analysis of current linguistic theories in the light of DNA genealogy data demonstrates that:
— the
Anatolian theory is generally compatible with DNA genealogy data; — the Vasconic and Afro-asiatic
substratum theory is partially in agreement with DNA genealogy data; — the Kurgan theory and the Palaeolithic Continuity Theory (PCT) appear incompatible with the history of Europe based on
haplogroup data. — the "Out of Africa" theory has questionable validity.
Keywords: Y Chromosome; Mutations; Haplotypes; Haplogroups; SNP;
Linguistics; Anatolian Theory; Vasconic; Kurgan; Palaeolithic Continuity
Introduction
DNA genealogy is an historical science that allows researchers to trace the
migration and evolution of populations. DNA genealogy studies the molecular history of DNA by
analyzing the mutations in the Y chromosome (in males) and in the mtDNA (in males and females). The
haplotypes of the Y chromosome are rather accurate tools; for example, using 111-marker
haplotypes resolves DNA-lineages down to 5-generation increments; mtDNA is a much cruder tool, and
its resolution stops at a few thousand years.
Many distinct linguistic theories have attempted to pin down when ancient
populations speaking different languages settled in Europe after the last Ice Age. These theories
offer supporting arguments, discuss the interrelationships with other theories, and often contest,
contradict, or reject aspects of other linguistic theories concerning the settlement of ancient
Europe. In this paper, we
1) establish a reasonable migration/linguistic/settlement paradigm for
ancient Europe from the Paleolithic to the Common Era, and
2) summarize the major linguistic
theories of the last 120 years. Then, using the tools of DNA genealogy, we
3) compare our results
with the hypotheses of the linguistic theories.
Ancient Migrations to, from, and within Europe as Revealed by DNA Genealogy
The α-haplogroup of the Y-chromosome (cf. Figure 1),
which is present in almost all males living today (except certain archaic African lineages A0, A00,
etc., not shown in Figure 1, since their dating is still uncertain), arose
around 160,000 ybp (Klyosov & Rozhanskii, 2012a) in a location currently unknown. Essentially, the
α-haplogroup was carried by the common ancestor of what we think of as anatomically modern
man. We can only conjecture where that common ancestor might have lived; it seems that he could have
lived in the vast triangle from Central Europe and Ireland to the west, through the Rus
sian Plain
to the east, to the Levant in the south (Klyosov & Rozhanskii, 2012a). This huge area is defined by
the greatest number of ancient skeletal fragments of anatomically modern homo sapiens (AMH) found in
Europe (dated between 45,000 - 43,000 ybp) (Benazzi et al., 2011; Higham et al., 2011), and in the
Russian Plains of eastern Europe (dated between 40,000 and 35,000 ybp or even between 45,000 and 42,000 ybp, when op
tically stimulated
luminescence dating of the settlements is used) (Prat et al., 2011; Anikovich et al., 2007).
Figure 1. Haplogroup tree of the H. sapiens Y-chromosome derived from
haplotypes and subclades (Klyosov & Rozhanskii, 2012a). The African branch is on the left, the
non-African one is on the right. The diagram was composed using 7415 haplotypes from 46 subclades of
17 major haplogroups. The timescale on the vertical axis shows thousands of years from the common
ancestors of the haplogroups and subclades.
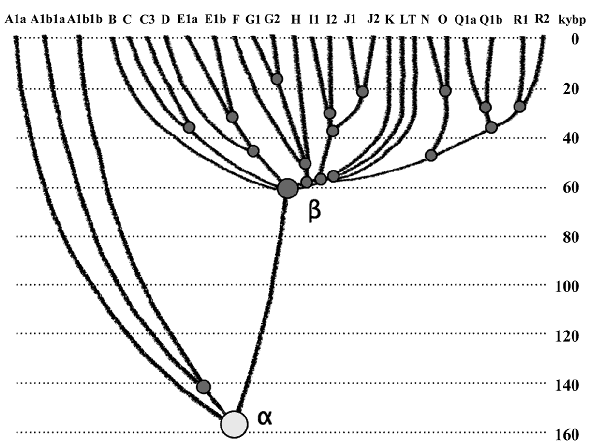
Figure 1 shows the estimated dates of the occurrence of human
haplogroups (Klyosov & Rozhanskii, 2012a). To compose this tree, we analyzed 7415 haplotypes from 46
subclades of 17 major haplogroups. The
α-haplogroup, which is ancestral to both the African and non-African haplogroups, arose about
160,000 ybp. The left branch represents current African haplogroups, which arose 160,000 - 140,000
ybp. The non-African
β-branch arose ~64,000 ybp; β and its descendants were not descendants of the
African branch but share a common ancestor. Haplogroups F through T represent Europeoids
(Caucasoids) who arose ~58,000 ybp (Klyosov & Rozhanskii, 2012a).
Some contemporary Africans are bearers of recently discov
ered haplogroups A0,
A00, etc. which arose some 200,000 - 260,000 years ybp, or even earlier (Mendez et al., 2013). These
might be the only truly African haplogroups. With respect to mutation, they are very distant from
other haplogroup A haplo
types.
This study concentrates on the haplogroups on the right-hand side of the diagram
in Figure 1. Two of them, R1a and R1b, descended from the R1 haplogroup, which
is shown in
Figure 1. R1a is the group, conditionally called the Aryans, which em
braces about
50% of the current population of Eastern Europe. This group has the same DNA as the legendary
Aryans, who arrived to India around 3500 ybp. Currently, approximately 72% of the some upper Indian
castes belong to the R1a haplogroup (Sharma et al., 2009).
Haplogroup R1a apparently arose about 20,000 ybp (Klyosov & Rozhanskii, 2012b) in
central Asia and possibly in the southern Siberia region of the Altai Mountains. Its ancient sub-clade M17 is observed in north China (Klyosov, 2009). R1a bearers migrated from central Asia across
Tibet, Hindustan, the Iranian Plateau, and Anatolia between 12,000 and 10,000 ybp. Their downstream
subclade, M417, crossed Asia Minor and entered the Balkans between 10,000 and 8000 ybp. It is
apparently their arrival in the Balkans which strontium isotope measurements dated at 8200 ybp
(Boric & Price, 2013). The M417 subclade spread all over Europe sometime between 9000 and 5000 ybp.
Around 5700 ybp, the recently discovered Z645 branch of haplogroup R1a developed. In 4900 ybp
(Rozhanskii & Klyosov, 2012), we find a Eurasian branch, Z283, and its South-Eastern branch Z93,
along with the downstream branch Z342.2/Z94 and the central Eurasian branch Z280. The central
Eurasian branch R1a-Z280 embraces about half of all contemporary east European males, and the
Aryan branch R1a-Z94 is currently observed in Russians, Ukrainians, and in southern Asian
populations in like the Kyrgyz, the Kazakh, and the Tajik peoples. This branch also exists in Iran,
India, in the Middle East, and along the ancient migration route from the Russian Plain to the
Middle East, particularly in Armenia and Turkey. The R1a haplotypes which were excavated in the
Andronovo archaeological sites east of the Ural Mountains, and which have been dated at between 3800
and 3400 ybp (Keyser et al., 2009), probably belong to the Z94-L657 subclade (Klyosov, 2013).
It seems that only two subclades, Z94 and L657, can be considered descendants
of the Aryans in the traditional sense. These subclades match the history, archaeology, and
languages of the steppe people. They rode chariots and, in the middle of the 2nd millennium BC,
arrived in India (Indo-Aryans), Iran (Avesta Aryans), and Mesopotamia (Mitanni Aryans) (Klyosov &
Rozhanskii, 2012b).
R1b bearers, called the Arbins, comprise about 60% of the current population of
western and central Europe. R1b apparently arose around 16,000 ybp (Klyosov, 2012b) in central
Asia, perhaps in the Altai region. Its subclade, M73, is ob
served in Siberia and central Asia;
subclade M269 is found in Bashkortostan near the South Urals; between 6200 and 5500 ybp, subclade
L23 and its downstream subclade Z2105 can be found on the Russian Plain, in the Caucasus, and in
Mesopotamia; between 5500 and 5000 ybp subclades L51 and L11 are found on the migration route
between the Middle East and the Pyrenees.
1022
R1b-U106 and P312 arose in Iberia around 4800 ybp and apparently became the initial population of the Bell Beaker culture of
continental Europe. L21 apparently arose in the south of France about 4000 ybp and moved to England
and Ireland sometime later. A common ancestor of nearly 25% of the current Irish population, who
lived around 1500 ybp (Klyosov, 2012b), belonged to M222, a subclade downstream of L21.
In addition to R1a bearers, since ~9000 ybp in Europe, and R1b bearers, since
~4800 ybp in Europe, ancient Europe was inhabited by bearers of other haplogroups, among them E1b,
G2a, F, I1, I2, J2, K. Their migration routes and dates of arrival in Europe remain obscure.
Haplogroups E1b and J2 apparently moved to Europe from North Africa or from the Middle East.
Haplogroup G2a moved apparently from the Near Asia, probably from the Iranian Plateau. Haplogroups
I1 and I2 might have moved westward from the Russian Plain, as had haplogroups IJK (see
Figure 1), between 45,000 and 40,000 ybp. The arrival of haplogroups F and K in Europe
has not been dated.
Recently, ancient bones in Spain dated as 7000 ybp (Lacan et al., 2011) have been
shown to belong to E1b-V13. Strikingly enough, present day bearers of E1b-V13 haplotypes all
coalesce to a common ancestor who lived only 3600 ybp. In other words, the contemporary V13
haplotypes reveal a gap between 7000 and 3600 ybp. The same gap pattern is observed in almost all
the haplotypes of ancient Europe — except haplogroup R1b, which apparently played an important role
not only in the settlement of, but also in the replacement of other haplogroups in Old Europe.
It seems that the arrival of the Aryans (R1a) in Europe had been peaceful; there
are no indications that it might have been genetically or otherwise violent. However, the arrival of
the Arbins (R1b) was marked by almost complete elimination of the autochthonous haplogroups from
Europe; E1b-V13 practically disappeared, and started to proliferate only around 3600 ybp; G2a fled
to the Asia Minor and to Mesopotamia and Caucasus; R1a fled to the Russian Plain; I1 nearly
disappeared and started to proliferate only around 3600 ybp; I2 fled to England, Ireland, and to the
Russian Plain. I2 started to proliferate in Eastern Europe only around 2300 ybp. Only R1b itself has
proliferated without pause from approximately 4200 ybp; a gap between 4800 and 4200 ybp is not
filled with their common ancestors as yet.
This brief historical outline of the settlement of Europe pro
vides the basis
for our consideration of the movement of peoples and languages in Europe from about 9000 to the
beginning of the Common Era (2000 ybp).
How Haplogroups and Languages Are Connected
DNA genealogy allows us to trace the migrations of ancient tribes and peoples, but, to date, it
has not helped us to unambiguously trace languages. Neither haplogroups nor languages stay the
same in the course of migrations: haplogroups can disappear as a result of extermination, epidemics,
and ecological catastrophes; in such cases the languages spoken by the haplogroup bearers
typically disappear. On occasion, however, the languages are adopted by other tribes. In some cases
the invaders adopt the language of the conquered people — when, for example, the women of a conquered
people continue to teach their own language to their children, or when a conquered people has a more
advanced civilization than its conquerors.
| The thesis of an advanced civilization imposing its language on backward peoples is a standing
mantra of the eurocentrism, it is self-serving and flawed. It has some factual support, but that
support is based not on the sophistication and benevolence of the advanced civilization, but on the
advanced greed, violent imposition and violent subjugation. The influence of advanced civilization
manifests in cultural borrowings, not in a language replacement, which invariably has a barbaric
mechanism, even if it is dressed in civilizing garments. If it is true that mothers of the conquered
people (and of the conqueror people alike) teach their language to their children, then the “advancement”
of the conqueror is irrelevant. And it is true not only for the mammals, but for the
dinosaur-descended birds too, all pups and chicks speak the language of their mothers. English is a
good example of that, while the Latin became a second language and a lingua franca of the privileged
few, it did not percolate the masses, who retained their own version of the English, and keep
surprising etymologists with folk lexus appearing from nowhere. In such cases the trend is the
opposite, it is the privileged few who pick up and start using the native language of their people.
This inaccurate premise is irrelevant to the contents and conclusions of the article. |
Even if the haplogroup maintains itself during the course of long migrations,
languages evolve following the rules of glottochronology and the natural dynamics of linguistic
evolution.
| The appeal to glottochronology and the natural dynamics probably is a lip service. Since 1492,
hundreds of languages have evolved, none of them predictable by the “rules of glottochronology and
the natural dynamics of linguistic evolution”, at least not the rules that have been developed so
far. The inability to forecast is an innate property of the old linguistics, and the forecasting
linguistics has not been born yet. Languages are developed by unpredictable stochastic processes. No
rules could have predicted that the Slavic language would live without significant changes through
250 years of Mongolian colonization, but would undergo a momentous transformation in the first 100
years after the fall of the Kipchak Khanate fragments that turned the Slavic into the Eastern
European Russian language. |
However, in some cases, a language can migrate and evolve along with the
migration and evolution of haplogroups over long periods of time and over large distances. There are
several conditions that must be met if our study of these cases is to be productive:
1) the
connection between the haplogroups and languages has to be verified by linguistics, DNA, and archaeology,
2) the languages must have evolved in time and distance,
3) the languages can be adopted by bearers
of different haplogroups in certain cases.
We have said above that haplogroup R1a migrated across Anatolia to the Balkans
between 10,000 and 8000 ybp; the group spread throughout Europe, moved east to the Russian Plain,
and then went to India. The first date is supported by the fact that we find PIE in Anatolia between
10,000 and 9000 ybp (Gray & Atkinson, 2003; Bouckaert et al., 2012). PIE could have been formed
and evolved during the long migration from the Altai Mountains to Anatolia. Then, the language
migrated with the same R1a haplogroup to the Balkans and across Europe, where around 6000 ybp it
split into branches; members of haplogroup R1a arrived around 4800 - 4600 ybp on the Russian Plain as
speakers of Indo-European language(s). DNA genealogy has confirmed that haplogroup R1a arrived in
India as the legendary Aryans around 3500 ybp; even today nearly 72% of some Indian upper castes are
R1a bearers (Sharma et al., 2009).
| The fact of PIE in Anatolia between 10,000 and 9000 ybp refers to a fact that some linguists
published their inferences. There is no material evidence of the truth or untruth of the inferences
made in the publications. No inscriptions were found, since the alphabetic writing has not been
invented yet till the 4000 ybp, or for another 5,000 years. Unlike the discovery of Neptune at the
tip of the pen, confirmed immediately after its location was predicted, the assertion of penning
linguists would remain an unprovable speculation, and is treated as such by less-enthusiastic
linguists. No PIE at 10,000 ybp, no migration, no split, no arrival in the Eastern Europe. What is
left? The R1a arrived in India around 3500 ybp. Arrived as the gold dust in the stream of other
haplogroups, unless it can be shown that by some mechanism the dust stratified from the stream and
appeared in a a proud solitude. Can it be shown? We do nor know till we see it. What we do
positively know, is that the stream did not bring over to India a Kurgan culture, lactose tolerance,
light hair and eyes, or a lexicon of the mounted horse riding. Until the opposite is proven, the
glorified migrants came in carts and brought with them PIE lexicon. |
Therefore, it seems that it was indeed haplogroup R1a carried PIE from about
20,000 to 10,000 ybp, and IE (or some kind of proto- or pre-IE languages from about 10,000 to 3500
ybp. The facts that
1) the peoples of the Russian Plain continue to speak IE languages, and
2) up to
63% of Russians today belong to haplogroup R1a, and
3) there are marked similarities between the
Slavic languages and Sanskrit, permit us to conclude that the migration of bearers of haplogroup R1a
were also bearers of Proto Indo European and Indo European languages.
The link R1a - IE does not hold the water:
1. The people of Australia and Siberia continue to speak IE languages, so
what? Slavic penetrated Eastern Europe in 600s, absorbed native and migrant Finno-Ugrian and Türkic
populations, and still continues doing that; the cultural change is unrelated to the Y-DNA marker.
English penetrated Australia in 1800s and replaced native language and native DNA marker, so it is R1b and IE. Siberia
was colonized after 1700s, largely replacing and supplanting native languages and wiping out native DNA,
so now it is a vinaigrette of DNA and IE. It can be
accurately stated that Eastern Europe, Australia, and Siberia continue to speak IE languages, but
that has nothing to do with the R1a haplogroup.
2. “Russians”
is a politonym, it embraces Slavic-Russians, Türkic-Russians, Chinese-Russians, etc. There is no
proof and no reason to assert that the original Slavic migrants were predominantly R1a-type. Across
the Slavic lands, I-type is most prominent, suggesting that the original Slavic settlers were
predominantly I Y-DNA. Before
the emergence of the Old Rus, the Eastern European population was Finno-Ugric and Türkic-Kipchak, Türkic-Bosnyak and Oguz,
Türkic-Khazarian, Türkic-Bulgarian, Türkic-Avarian, Türkic-Hunnic, and now they are
all Russians. Most of the Türkic people belonged to the Ogur branch and had R1a Y-DNA marker.
Culturally Russian, genetically they are R1a-type Kipchaks, Bosnyaks, Oguzes, Khazars, Bulgars,
Avars, Huns, and many more, constituting most of the 63% Russian R1a demographics. The Slavs, who
amalgamated with the Türkic Sarmatians and Huns before 600s, also had some R1a.
3. To explain similarity between the Slavic and Sanskrit languages, is good enough to look at the
situation before the Aryan out-migrationto from the Eastern Europe to the South-Central Asia
4000 ybp, and at the Corded Ware Culture. No reason to ascend to the putative 10,000 ybp, and much less
so to the dim 20,000 ybp. |
We can add to our earlier description of haplogroup R1b’s (the Arbin’s) migratory
route the following points: around 6500 - 6000 ybp, on its way from the Russian Plain south over the
Caucasus and probably — concurrently — along the eastern side of the Caspian Sea and Eastern Iran, it
moved to the Middle East, the Tigris and Euphrates basin; between 6000 and 5000 ybp it apparently
established the Sumerian civilization; between 4800 and 4500 ybp it moved to Europe following
several routes. One route brought the Arbins through Northern Africa to the Pyrenees. Between 4800
and 4500 ybp, they arrived in continental Europe as bearers of the Bell-Beaker culture; another
route brought the Arbins to Europe through the Mediterranean islands and the Apennines; around 4500
ybp, yet another route brought the Arbins to Europe via the Pontic steppes.
In the first part of their migration, along the northern Eurasian route, the
Arbins crossed territories, populated at least for the last two millennia (and very probably also
much earlier), by speakers of Turkic languages, such as Chuvashes, Bashkirs, Tatars. We can conclude
that the Arbins might have carried languages which were proto-Turkic, or Dene-Caucasian, or
Sino-Tibetan. We tentatively call these languages Arbin, or, or Non-Indo-European (NIE)
agglutinative languages. In the Caucasus, the Arbins left the northern Caucasian group of languages, together with a characteristic vigesimal counting system. Two thousand years later, the Arbins brought the same base-20 counting system to the Pyrenees.
| Genetically, references to the Dene-Caucasian and Sino-Tibetan languages appear to be a preemptive
lip service. The R1b migration history and R1b NIE in Europe is unrelated to the Q Y-DNA Dene-Caucasian
and O Y-DNA Sino-Tibetan. |
103
The R1b bearers brought their Arbin language(s) first to Mesopotamia, then to the Sumer state
(Assyrians, the likely descendants of the Sumerians, today are largely R1b bearers, which is unusual
for the Middle East [Klyosov, 2012b]), then to Iberia, where the present day Basques, 87% - 93% of
whom belong to haplogroup R1b, also employ the vigesimal counting system. As Bell Beaker tribes
the Arbins moved north to continental Europe, and brought their agglutinative NIE languages, which
apparently were spoken in Europe between 4500 and 3500 - 3000 ybp, and up into the Common Era (e.g.,
probably, Picts) and to the present (Basques).
| Genogeography keeps asserting that Basques are Iberian natives because they share alleles with other
Iberians. Testing mtDNA would not produce different results because for millennia, the mamas were
locals, passing on their local make-up. Import is recorded in Y-DNA. |
During the period of 3000 - 2300 ybp many R1a tribes migrated with their IE
languages from the Russian Plain to central, western and southern Europe bringing to Europe the
peoples later called Germans, Italics, Greeks, Illyrians, Balto-Slavs, and Celts (the Hallstatt and
La Tene cultures flourished between 2600 and 2400 ybp). We posit that some Arbin peoples adopted the
IE languages from the R1a bearers and, in exchange, introduced NIE loan words and grammatical
structures. One group of Arbins were forebears of the Basques in the Pyrenees and the South of
France, as well as the Picts in northern Scotland, and, possibly, the Etruscans in Tuscany.
Linguistic Theories Regarding the Ancient European Settlements
Let us move now to current linguistic theories about ancient European
settlements, and compare their notions with those of DNA genealogy.
The Vasconic and Afro-Asiatic Substratum Theory: The Linguistic View
The Vasconic and Afro-asiatic substratum theory of Vennemann (2003) proposes
that several millennia after the end of the last Ice Age, when the glaciers receded (around 10,000
ybp), NIE peoples started to settle in southern Europe. These peoples were responsible for many
European toponyms, hydronyms, and floral and faunal names, some of which have survived up to our
times. Krahe (1954, 1964) believed that many of these toponyms and hydronyms were Indo European, but
Vennemann was convinced that they contained NIE roots. Krahe argued that hydronyms from the Atlantic
were imposed on the Baltic shore areas before 3500 ybp, and
preceded the formation of the IE Baltic, Celtic, Germanic, Illyrian, Venetic and
Italic language groups. Because of their similarities, Krahe concluded that the toponyms and
hydronyms descended from a common language system he named Old European (OE). According to him, Old
European constituted a language layer intermediate between PIE and the IE Baltic, Celtic, Germanic,
Illyrian, Venetic and Italic language groups.
Schmid (1987, 2001) extended Krahe’s OE concept by including the Eastern Slavic
languages. Vasconic is what Vennemann called the language family of the NIE populations which
imposed the toponyms and hydronyms. The Basque language would be the only surviving language of this
family. Another argument in support of the Vasconic theory is the persistence in modern languages
of traces of the base 20 counting system that would be a relic of the Vasconic culture.
Vennemann (2003) also observed that on the Atlantic shore area of Europe there
are toponyms that are neither Vasconic nor IE. He named the languages responsible of these toponyms
Semitidic. According to Vennemann, these languages were related to the Mediterranean
Hamito-Semitic languages, and were spoken along the Atlantic shore between 7000 and 3000 ybp. The
Semitidic languages influenced IE superstratically (i.e., loaning terms for animals, advanced cattle
breeding, buildings, warfare, and social organization — especially among the Germans of northern
Europe) and substratically (i.e. contributing loan terms for plants, animals and herding, especially
among the insular Celts). From about 7000 ybp onward, the Semitidic peoples — thought to be builders
of megaliths — moved north along the Atlantic coast, reaching Great Britain and Ireland about 6000 ybp
and Sweden about 5000 ybp.
According to Baldi et al. (2006) there are several weak points in this theory: no
megaliths have been dated before the Bronze Age (3500 - 2800 ybp); contrary to the traditionally
accepted evidence that the Celts settled the Britain and Ireland no earlier than 4000 ybp,
Vennemann’s theory requires a Celtic presence in England and Ireland about 7000 ybp; the building of
megaliths by Semitidic settlers is opposed by Renfrew and other archaeologists; finally,
Vennemann (2003) assumed that the Picts of northern Scotland were an Atlantic population or at least
a population speaking an Atlantic language. A similar hypothesis, according to which the Picts were
a NIE people, was set out by Zimmer (1898) on the basis of the Pictish customs of tattooing and
their matrilineal social organization.
Vennemann assumes no genetic connection between IE languages and Vasconic and
Semitidic languages. The expansion of the OE toward north Europe was restricted by the expansion of
IE populations which adopted the Vasconic toponyms, hydronyms, and other lexical items related to
the natural environment. The Basques, now living in a restricted region between France and
Spain, speak the only descendant language of OE or, according to Trask (1995, 1997), a patchwork of
NIE languages is uncertain.
Kuzmenko (2011) has reviewed the lexical borrowings made by the Indo-European
languages of Europe from an "unknown substrate." In his opinion, most linguists of the last century
agree that an unknown substrate contributed not only to German languages but to all European IE
languages. Kuzmenko finds merit in Vennemann’s hypothesis (2003) that the Basque language is the
only surviving representative of the unknown European substrate.
The Vasconic and Afro-Asiatic Substratum Theory: The View of DNA Genealogy
The Vasconic and Afro-asiatic substratum
(VAAS)
theory is partially confirmed by DNA genealogy.
DNA genealogy does not support the assumption of the VAAS theory that NIE
populations began their European settlement in southern Europe after the end of the last Ice Age
(about 10,000 ybp). Instead, it reveals that between 4800 and 4500 ybp (Klyosov, 2012b) the Arbins
(R1b) moved into Europe using several routes (Northern Africa and the Pyrenees; the Mediterranean
and the Apennines; the Pontic steppes). There were no speakers of Vasconic in Europe before 4800
ybp.
| Dating and routing define expected technological and biological developments. The
circum-Mediterranean route started in North Pontic at 6500 - 6000 ybp, carrying Mesolithic technology,
hunter-gatherer traditions with incipient animal husbandry economy, Tengriism with early Kurgan
tradition, and Mesolithic form of language. On the way that lasted 1200 - 1700 years, or 48 - 68
generation, the early Kurgans had to adopt new technologies, new allies, E and J Y-DNA haplogroups,
and considerably divergent language influenced by agglutinative Dravidian and flexive Hamito-Semitic
(Afroasiatic) languages, especially with the cultural terms. The demographical bottleneck forced an
evolution in the circum-Mediterranean migrants, amplifying some survival skills, obliterating the
others, and possibly changing their allies and genetic composition. The Pontic steppes overland route is
depicted as Kurgan three waves, with early and developed animal husbandry economy, high mobility, Chalcolithic
technology, Tengriism with strong Kurgan tradition, and compared with circum-Mediterranean migrants
later forms of language. The waves were not identical nor uniform in their genetic make-up, joined
by different allies of N and other haplogroups, and in Europe enriched by I and R1a Y-DNA. The waves
are connected with the eastward and westward spread of Seima-Turbino metallurgy. |
104
However, the notion that Vasconic is a descendant of the ancient Arbin language
is in agreement with DNA genealogy data. Concerning the European toponyms, hydronyms, and the names
of flora and fauna which have survived to the present, Venneman’s hypothesis — that they are NIE — is
acceptable, provided that his temporal estimate (10,000 ybp) be adjusted to 4800 ybp or later. DNA
genealogy is in general agreement with the hypothesis of Krahe — that the languages are OE
(Old Europe) because,
according to DNA genealogy, the Arbins (R1b) and their NIE languages migrated as bearers of the
Bell-Beaker culture (mainly R1b) and apparently dominated Europe between 4500 and 3000 ybp. Krahe
(1954, 1964) appears to be correct in assuming that the Vasconic toponyms and hydronyms were imposed
before 3500 ybp.
According to DNA genealogy, the Vasconic language family is nothing other than an
alternate name for the NIE languages of the Arbins (R1b). In other words, the NIE language of the
contemporary Basques (R1b haplogroup) is probably a surviving descendent language of the NIE
languages of the ancient Arbins (R1b). A common ancestor of present day Basques, most of whom belong
to haplogroup R1b, lived around 3700 ybp, which reflects a population bottleneck of the Arbins who
arrived in Europe 4800 years ago (Klyosov, 2012b).
The vigesimal counting system used both by the Basques and by the people of the
Caucasus is supported by DNA genealogy data as a characteristic suggesting a connection between the
ancient Arbins (R1b) (who migrated along the Northern route across the Caucasus, the Mediterranean
islands, and northern Africa to Central Europe) and the isolated ancestors of the Basques in the
Pyrenees region.
Concerning the Semitidic, or Atlantic group of languages postulated by Vennemann
(2003), they might indeed have survived into the Common Era, and could have been spoken by the
Picts of northern Scotland. The haplogroup of the Picts is unknown at present, but it might have
been I1 or I2, because both haplogroups can be found in Britain today, and their common ancestor
lived more than 15,000 ybp (Klyosov, 2010; see also
Figure 1). The majority of the population of England and Ireland carry the R1b
haplogroup, which came to Britain and Ireland after 4200 ybp. Indo European apparently belonged to
the Aryan tribes (R1a); Non Indo European belonged to the Arbin tribes; Semitidic belonged to
haplotypes I1, I2, and G2. The three linguistic communities had a common ancestor who lived around
55,000 ybp (Klyosov & Rozhanskii, 2012a). Therefore, Vennemann’s suggestion that there were no
genetic connections between IE languages, Vasconic languages, and Semitidic/Atlantic languages
seems to be justified.
| The conical hats of the Picts, their tattoo tradition, and small ungulate economy make them
oddball in Europe. The closest in time parallels are located in the Middle Asia. The Y-DNA
R1b-L21/S145, aka Scots Modal haplogroup, is most common in the area with most dense Pictish
symbols. The subclade L21 split from the circum-Mediterranean Y-DNA R1b at about 4000 ybp, attesting
that Scotts and Picts (both late Latin monikers) ascend to the N. Pontic - Iberia
circum-Mediterranean migrants. The subclades of the migrants marked by M269 subclade became known as
the Bell Beakers and populated Europe from the Atlantic seaboard eastward to the Balkans, Carpathian
Mountains, present day Poland to the Baltic Sea and to the western border of the East European
Plain. Another trait of the Scottish and Irish people, their attention to genealogy, is attested
among the
Türkic people, who had to memorize their ancestry for at least 7 generations, and who monitored
tribal relationship for millennia, to prevent insest. Given a choice between I and R1a haplogroups
as markers of the IE language, the haplogroup I has numerous advantages: it was the most frequent
European haplogroup before the almost complete elimination of the E1b, F, G2a, J, I1, I2, and K
haplogroups from Europe after the calamity of the “killing fields” 5,000 ybp; it was the only
haplogroup that could endow Scandinavia with their version of the IE language, the other component
being NIE R1b, it is the predominant haplogroup of the Central European Slavs, whose language is
closest to the Sanskrit; it is the haplogroup that survived in Scandinavia and Eastern Europe the
period of the “killing fields”, and re-populated Europe again. Against the I Y-DNA, R1a Y-DNA has
all disadvantages: it populates the historical territories of the
Türkic Ogur tribes in modern Russia; it could not be brought over to Russia from the
I-dominated Central European Slavs; the Boyar and Brahman castes in India are likely descendents of
the Huna and their kindred tribes that retained socially dominant status by switching to the local
languages; its initial origin, with NIE R1b, from a common genetical trunk points to its originally
NIE linguistic affiliation; and paleogenetics found R1a among the NIE Eastern Hunnic elite. |
The Anatolian Theory: The Linguistic View
Renfrew (2001), in summarizing the Anatolian theory, affirms that PIE, or the
PIE family of languages, or the pre-PIE languages (Diakonov, 1984), were formed in central Anatolia
during the Neolithic (about 9000 ybp), and that the PIE or IE languages were diffused throughout
Europe from West Anatolia along with the diffusion of the agriculture, which was Phase I of the
PIE.
According to Renfrew (2001), reliable radiocarbon datings indicate that the
domestication of plants and animals from West Anatolia reached Greece and Crete around 8500 ybp.
Linguistic changes in Greece and in the Danube and Balkan areas were due mainly to demic migrations
during the 9th and 7th millennia ybp. It is possible that around 3500 ybp the diffusion of agriculture east of what is now Ukraine could have brought speakers of Tocharian to the Chinese
Sinkiang/Xinjiang.
On the basis of their study of 87 languages and 2449 lexical items, Gray and
Atkinson (2003) and Gray et al. (2011), suggest that an initial IE divergence occurred between
11,800 and 9800 ybp, allegedly in Anatolia. This is consistent with the separation of archaic PIE
from pre-PIE; Ryder and Nicholls (2011) indicate a unimodal posterior distribution for PIE at about
10,400 ybp, which supports the Anatolian theory; other linguistic studies by Sturtevant (1962),
Dolgopolsky (1987, 1993), Gamkrelidze and Ivanov (1984, 1995), Pringle (2012) and Bouckaert et al.
(2012) also support the Anatolian theory. Interestingly, Bouckaert’s study is based on a model of
spatial diffusion of infectious diseases. Renfrew (2001) affirms that a first linguistic advergence
area was formed in the Balkan region between 7000 and 5000 ybp, which was Phase II of PIE.
Some linguistic characteristics of the Celtic and Tocharian languages indicate
that they were not part of the Balkan linguistic advergence area. The disaggregation of the Balkan
advergence linguistic area, which occurred at around 5000 ybp, indicates the end of Phase II of
PIE, and the separation of proto-Greek from proto-Thracian, proto-Dacian, proto-Phrygian and others.
At about the same time, there was a separation of the proto-Indo-Iranian spoken in the northern area
of the Black Sea from its diffused form on the Iranian plateau and in India. Renfrew (2001) asserts
that other IE languages were developed in advergence areas, where now their descendant languages are
spoken.
The Anatolian Theory: The View of DNA Genealogy
The Anatolian theory is generally compatible with DNA genealogy data, although
the linguistic theory is silent about the evolution of PIE before 10,000 - 9000 ybp.
As we discussed above, the proto-Aryans (R1a) migrated westward across Anatolia
around 10,000 - 9000 ybp, which fits the linguistic estimates of Renfrew — 9000 ybp (2001), Diakonov — 11,800 to 9800 ybp (1984), and Gray et al. (2003, 2011). Diffusion of agriculture, demic
diffusion, and non-demic diffusion are concepts beyond the purview of DNA genealogy, though
migrations of the proto-Aryans (R1a) from Anatolia to the Balkans about 9000 - 8000 ybp could
represent Phase I of PIE. The later spreading of the Aryans (R1a) along with their IE languages
across Europe about 8000 - 5000 ybp could represent Phase II of PIE. The migrations eastward of
Proto-Aryans to the Russian Plain and their split (about 4500 - 3500 ybp) into at least four
migration routes to the south, southeast, east southeast, and east toward India could represent
Phase III of PIE.
| The revised 2004 version of the Anatolian Theory dropped dating by 2 millennia, now it stands at
9,000 ybp, and the PIE moved to Balkans and suggested 7,000 ybp. It is still far from the majority
Indo-Europeanists' opinion of PIE between 6,500 and 4,500 ybp, and the most probable date at 5,700
ybp. The model stipulates a common language (PIE) diversified into branches, it is a static model
without dynamic and stochastic conversion, and it is even not a Sprachbund. The late PIE predates
4500 ybp, and Proto-Indo-Iranian predates 4000 ybp. In Starostin version, the earliest PIE is
dated to 6,670 ybp, just for Tocharian, and the later PIE, for all other IE languages to 5,810 ybp.
These various later versions of the Anatolian Theory conflict with the view of DNA Genealogy much
more than the original much criticized and discarded Renfrew version.
The focus on the Tocharian languages is popular with IE linguistics, but is displaced. First, it
is widely recognized that the name is a politicized misnomer, the true names are Kuchean or Agnean
(Toch. A) and Karashar (Toch. B). Secondly, the dating has to be driven by Kucha, Agni, and Karashar
archeology, and not glottochronology that uses a circular logics. Thirdly, since the genetics
identified the Indian ancestry of the Tarim basin sedentary population, attention should shift to
the Indian IE dialects, still a white spot on the IE map. Forthly, the Kuchean or Agnean (Toch. A)
language was physically brought to the to the Meroites in the Kingdom of Kush in Sudan, together
with their Kharosthi script. The Kushana language of the Kushan Empire in India, and of the Kush
Kingdom in Sudan, grossly misnamed Tocharian, was an Indian lingua franca or trade language in
Central Asia used by diverse peoples living in an intense bilingual environment. The Kush Kingdom
left an abundance of Kushite documents written in Egyptian language before the introduction of
Meroitic language. The history of the Kuchean or Agnean (Toch. A) and Karashar (Toch. B) languages
is a page of the Kushan history.
The origin of the IE languages is confusing enough without using misnomers and false
attributions. |
The suggestion of the Anatolian hypothesis that Tocharian languages were not part
of the Balkan linguistic advergence area is conditionally supported by DNA genealogy. According to
Gray and Atkinson (2003), the Tocharian languages were an archaic branch, which arose around 7900
ybp, and were spoken by R1a populations in the Tarim basin. Based on the dating of the Tocharian
language and the relatively high linguistic distance of Tocharian A and B from the other IE
languages (Tomezzoli & Kreutz, 2011), it is unlikely that the proto-Tocharians migrated westward to
Europe and the Russian Plain with the proto-Aryans (R1a), and then
moved back to the Tarim Basin. It is more likely that the proto-Tocharians migrated
from the Altai region of north China to the nearby Tarim basin and remained there (never going to
Europe), forming the autochthonous R1a peoples of Central Asia. The Anatolian hypothesis groups
these Tocharians rather superficially with Europeans (Li et al., 2010), without any DNA
justification — their haplotypes were not even reported for a comparison with European R1a haplotypes. It is not
enough to consider Tocharians as Europeans on the basis of their somatic features and their
clothing which, in 4000 ybp, looked like Scottish plaid. In fact, plaiding techniques could equally
well have been brought to Europe by R1a tribes from the Altai and Central Asia.
105
Still, there is some room for the Tocharian languages to be considered as
derivatives of the archaic European R1a languages of the IE family. Tocharian is possibly an
ancient Centum branch. In that case, we have to admit that Gray and Atkinson’s (2003) estimate
of their appearance (7900 ybp) should be reduced at least to around 6000 ybp. There should also be a
recognition of an earlier migration (between 6000 - 5500 ybp) of R1a bearers from Europe to the
Altai region, and their possible contributions to the Afanasyevo archaeological culture and
perhaps to the Centum Tocharian languages in the area, including the Tarim basin. This concept is
verifiable; if Afanasyevo bones not too far away from the Tarim basin are dated at least 5000 ybp
and are shown to belong to the R1a-Z93 subclade, the case for a migration of R1a from Europe to the
Tarim basin will be well supported.
DNA genealogy data disallows Anatolia as the homeland of PIE and IE languages.
DNA records show that these languages had no specific homelands — R1a bearers migrated over thousands of miles during the course of thousands of years. No archaeological site can be possibly
identified as a location in which IE split into branches — the branching of IE was a continuous
process of divergence and convergence over millennia.
| The thesis on divergence is a cornerstone of all presented IE theories, but where the convergence
came from? It stands outside of these theories, it opens a door to uncounted complications, and it
brings aspect of stochasticity into otherwise linear world. The orderly retroactive application of
the linguistic laws goes out of window, and the proto-language needs to be redefined. In a
convergence, a parent language of a proto-language is not a proto-language by definition, it is two
or more amalgamated substrates, like O + H2 > water, a new substance. If a linear model
of Y-DNA may be congruent with a linear linguistic model, it can't be congruent with a stochastic
linguistic model, and vice-versa. Something must give in, either a linear Y-DNA goes out of window,
or a linear linguistic model goes out of window, or both. In real life, both Y-DNA's conglomerate,
and languages amalgamate. |
According to DNA genealogy data (see
Figure 1), the predecessors of those who spoke PIE languages might have migrated
50,000 ybp or earlier from the unknown birthplace of the β-haplogroup. The birthplace
might have been in Europe, the Russian Plain, or south Siberia (where they arrived between 40,000 -
35,000 ybp). Much later, sometime after 20,000 ybp, they migrated westward along with the R1a
haplogroup via Anatolia, to the Balkans, to the Russian Plain and Pontic steppes, to the Middle
East, Middle Asia, the Iranian plateau, the Ural mountains, Hindustan, South Siberia (again), North
China, and Mongolia. All of these locations are migrational passing points and not homelands for the
predecessors of the IE languages.
| Except for few island languages untouched by outside influences, all modern languages are
amalgamations, are distinct languages, and are proto-languages of future languages, like Spanglish
and Russinitic. All distinct languages had a distinct territory, and those territories are called
homeland. Some homelands were for a short stay, some more stationary, but they all laid on the
migration routes. Dismissing linguistic homelands as mere passing points throws away baby with the
water. The transitory nature of the homeland is just another property of the linguistic process, an
objective reality independent of scientific fantasies. |
The Kurgan Theory: The Linguistic View
During the Mesolithic and the Neolithic, during the dry and cold period of the
Younger Dryas (12,800 - 11,500 ybp), NIE and PIE peoples settled along the shores of the Black Sea.
According to the Kurgan theory, Proto Indo European formed in this area. At about 7600 ybp (Ryan
et al., 1998), due probably to a cataclysm, the waters of the Mediterranean Sea entered the Black
Sea through the Bosporus, triggering a rise in the water level and the submersion of many human
settlements. The cataclysm caused extensive migrations toward the Balkan region and Central
Europe; it ultimately gave rise to the formation of the Neolithic cultures of Vinča
and the Linearbandkeramic (LBK).
| 12,800 - 11,500 ybp probably all languages were NIE, unless the PIE definition is stretched to
embrace any stage in linguistic development. Example: (reconstructed) PIE küjna (dog,
cannine), p'at (foot, pedicure), haku (water, aqua), and küni (woman, gynecology). (Reconstructed)
Proto-Amerindian: akuan (dog, cannine), pet (foot, pedicure), haku (water, aqua), and
kuna (woman, gynecology). The common origin is obvious, hence the Amerindian is PIE, the
Redskins are IE, the Q Y-DNA is IE, and the R1a PIE goes out of the window. However, if
agglutinative and non-flexive Amerindian languages do not fall under the definition of the PIE, they
are out, and R1a is back in business. To qualify them, all we have to do is to prove that R1a is
exclusively non-agglutinative and flexive. Like the adage says: law is a pole, where you turn there
you go. |
Marija Gimbutas (1991) defined Ancient Europe as the European Culture developed
between the 9th and the 7th millennia bp in the area of the Balkans, Greece, Adriatic region,
Moldavia and Ukraine before the arrival of the IE bearers (actually, in Gimbutas
they did not arrive, they are authochtonous). Earlier (1956), she had provided a
rather comprehensive description of the cultural level of an ancient Europe characterized by well
organized settlements, mixed (h)orticular economies, high quality sculpture and ceramics, and elaborate
religious traditions. This materialized in the cultures of Bükk, Butmir, Cucuteni-Trypillia,
Dimini, Karanovo, Lengyel, Petreşti, Vinča, and LBK.
The languages spoken in Ancient Europe were NIE, as indicated by the survival of NIE
agricultural, technological and social terms, toponyms, and personal and tribe names. Between 7500 and 6300 ybp
Ancient Europe developed an advanced civilization, excelling in metallurgy. The Model of the Steppe,
or the Kurgan model, or the Kurgan hypothesis, or the Kurgan theory was developed mainly by Gimbutas
(1994, 1997), (see the synthesis by Marler [2001]); it proposes the presence, in about the 7th
millennium bp, of territorial, nomadic, pastoral peoples speaking PIE languages. This Kurgan culture
was situated in the area of the Dnepr and Don basins, the middle and lower Volga basin, and in the
Caucasus and Ural mountains. The tombs, covered by round tumuli named kurgans often contained
weapons and other artifacts, suggest a culture that was patrilineal, pastoral (with rudimentary
agriculture), territorial, and nomadic. The Kurgans had domesticated the horse around 7000 ybp
(Bököny, 1997; Gimbutas, 1956).
The Kurgan culture had characteristics different from those of the cultures of
Ancient Europe, indicating to Gimbutas that it had not developed from the cultures of Ancient
Europe.
A first migration of Kurgan peoples, according to Gimbutas’ theory, took place
about 6400 - 6300 ybp, as a result of the progressive drying of the steppes during the 8th and the
7th millennium bp. The Kurgans moved towards Bulgaria, the Danube basin, and Central Europe. This
migration is given support by the increasing number of kurgan tombs (discovered between the
egalitarian tombs of the Ancient Europe cultures), the fortification of settlements, the damage to
the settlements of the Varna, Karanovo-Gulmeniţa, Vinča, Lengyel and LBK
cultures, and the replacement of some Ancient Europe cultures by new Kurgan cultures. The
development of IE languages was due to language substitution and bilinguism.
A second migration took place around 5500 ybp from the area north of the Black
Sea through Ukraine toward Poland, and central and east Germany. This migration led to the formation of hybrid-cultures: the Baden complex in the middle Danube basin (which had the Vinča
culture as substrate), the Ezero culture in Bulgaria (which had the Karanovo culture as
substrate), the Globular Amphora culture in Romania, West Ukraine, Poland, and Germany (which had
the Trichterbecherkultur [BK] as substrate). In parallel with the development of these
hybrid-cultures, the fragmentation of PIE into several IE languages took place.
| In M. Gimbutas linguistic hypothesis, PIE was born by stationary local people, specifically
Balts and Uralics (of N Y-DNA, as we know today), and branching occurred on the junctions of the PIE
and NIE languages. This is not a linear model, it does not fall under the Verner's law, Grimm's law,
and every other law concocted for the linear isle model, it is defying the schemes of the canonized
linguistics. The objective of the hypothesis is to bring the IE to Europe, thus the timeline starts
at conception and ends at delivery. After that, linguists can do what they please. Of all the IE Urheimat
theories, M. Gimbutas' theory is most popular. |
A third migration, this one from the Volga steppes, took place between 5000 and
4800 ybp. It was more massive than the other two as witnessed by the numerous Yamnaya culture
burials in the Balkan region and East Hungary. This migration caused the displacement of the hybrid
cultures of central Europe toward northern Europe, southern Scandinavia, the Baltic area, and
central Russia. This last migration was followed by a period of stability characterized by the
formation of cultural groups (Gimbutas, 1994, 1997) which spoke distinct IE languages.
The Kurgan Theory: The View of DNA Genealogy
DNA genealogy data suggests that the Kurgan theory is incompatible with the history of Europe.
106
According to the DNA data, PIE arrived in the Balkans after a long migration from
central Asia. Using strontium isotopic measurements, Boric and Price (2013) have shown a significant
increase in non-local individuals in the Balkans from ~8200 ybp. This generally coincides with the
arrival in the Balkans of R1a peoples and IE languages. Neither the people nor the languages came
from the Pontic steppes.
| Nothing attests to the arrival of PIE, or for that matter of any other linguistic category, at
~8200 ybp. The assertion is based on the modern predominance of R1a in the Brahman caste in India,
with a gap of 8,000 years between present and 8200 ybp, and totally unknown mechanism of the
emergence of the Brahman caste. An attested example of Luvians in Israel, where the IE-speaking
Luvians speaking archaic IE turned into Jewish Brahman caste of the Levites, the Luvians did not
retained their archaic IE lingo, but switched to the local Semitic vernacular. In Russia, the Greek
priestly caste turned into Slavic-speaking religious hierarchy. In Rome, the Aramaic-speaking
disciples produced generations of Greek- and Latin-speaking priestly caste. In the Muslim countries,
the descendents of the founder's clan switched from Arabic to local languages, still continuing the
castes of Seids. The assertion that Brahman's original language is the same as their modern
language, and not, like for anybody else, a legitimate result of social pressure, is quite daring.
The circular logics in this premise is obvious. First is postulated a linear Family Tree model, then
for that model is developed a glottochronology, the glottochronology suggests dates, and dates
confirm a linear model. |
DNA genealogy indicates
1) a migration of R1a peoples eastward from Europe to the
Russian Plain between 4800 and 4600 ybp (i.e. a direction opposite to that suggested by the Kurgan
theory), and
2) a migration of R1b peoples from Asia to the Russian Plain and then southward between
7000 and 5000 ybp, and westward, between 5500 and 4500 ybp.
In other words, the migrations of the Arbins (R1b) and the Aryans (R1a) were separated in time and went in largely opposite directions.
Overall, NIE speakers were moving west to Europe and south to the Caucasus; IE speakers were
moving east. The Kurgan theory posits language migration in the opposite directions. In other words,
the Kurgan theory distorts the whole pattern of what happened in Europe and the Russian Plain
between 5000 and 3000 ybp. Additionally, contrary to what is proposed by the Kurgan theory:
Expansion of Kurgan people
Map schematically depicts expansion in the Western Eurasia
In the Eastern Eurasia, Kurgan expansion reached Pacific
The Celtic Kurgan circum-mediterranean migration is not shown
The two arrows at the bottom right depict much later migration of non-Kurgan Indo-Aryan farmers.
Arrows show flow, not the waves, i.e. they are timeless.
https://en.wikipedia.org/wiki/File:IE_expansion.png
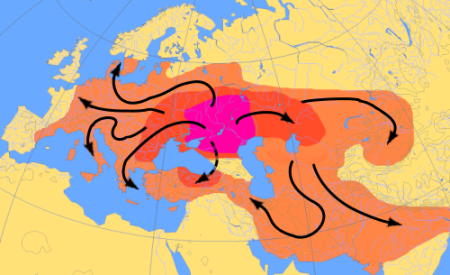 |
— PIE speakers and their languages were not settled or formed along the shores of
the Black Sea between 12,800 and 11,500 ybp. At present, we don’t know which haplogroups were the
most affected by the Black Sea cataclysm. The victims might have been G2a, E1b, F, I1, I2, etc.,
with survivors moving westward, to Europe.
— Ancient Europe cannot be considered as having an established European culture
developed between the 9th - 7th millennia bp in the area of the Balkans, Greece, the Adriatic,
Moldavia, or Ukraine before the arrival of IE. In fact, the IE speakers (R1a) arrived in the
Balkans and further in Europe between the 10th - 8th millennia bp.
— The languages spoken in Gimbutas’ Ancient Europe were not totally NIE. In fact,
the arriving IE speakers (R1a) introduced their IE languages between the 10th - 8th millennia bp.
The survival of NIE agricultural, technological and social terms, toponyms, and personal and tribe
names cannot be considered an argument supporting a totally NIE Ancient Europe, since from the 10th
millennium on IE and NIE languages co-existed in Europe
— Gimbutas’ theory is in error when it proposes the formation of territorial,
nomadic, pastoral populations speaking PIE languages (collectively named the Kurgan culture), in the
7th millennium bp in the area of the Dnepr and Don basins, the middle and lower Volga basin, the
Caucasus and the Ural mountains. In fact, there were no PIEs (R1a) at those times in those
territories. The Kurgan theory apparently has inverted the roles of the NIE (R1b) and the IE (R1a).
— The Kurgan theory is in error in ascribing kurgans, nomadism, and the
domestication of horses to speakers of IE who lived around 7000 ybp. Instead, these cultural
features should be ascribed to NIEs (R1b) who migrated westward.
— Gibutas claims that IE speakers migrated to Europe three times — first, between
6400 and 6300 ybp; second, around 5500 ybp (from the area North of the Black Sea); third, between
5000 and 4800 ybp (allegedly from the Volga steppes). These claims are unsupportable. There were no IEs (R1a) in the Volga steppes between 5000 and 4800 ybp or earlier; they arrived between 4600 and
4300 ybp. Had they been in the steppes, they would have been moving from Europe eastward. As we
suggested above, there might have been an ancient migration route for R1a bearers (between 6000 -
5000 ybp). That route has not been proven as yet. If it is proven, it will most certainly be a
migration to the east rather than the west as Gimbutas alleged.
| Seeing migrants as genetic blobs instead of a point would only reinforce the genetic
argumentation. The movements of the late migrations set realistic examples: the westward migration
of the nomadic Kurgan Oguzes from the Balkhash to the Aral area ca. 700 AD brought along a
constellation of tribes that aggregated with the constellation of the local tribes, forming a
24-tribe confederation. Each constituent tribe had its own lengthy history, marital partner, genetic
make-up, and vernacular. Oguzes have retained their
Türkic language, but it deviated a mile from their pre-migration language. The westward
migration of the nomadic Kurgan Bulgars ca. 700 AD brought along a constellation of tribes that
aggregated with the constellation of the local tribes, forming a new confederation. Each constituent
tribe had its own lengthy history, marital partner, genetic make-up, and vernacular. Bulgars mostly
lost their
Türkic languages, only a minority of them have retained them, and the modern Bulgaria uses
Slavic.
In these two examples that attest to only two models of very many observed, the migrants
genetically were of different colors, shades, and smells, and in the destination place the blobs
transformed and changed their colors and smells, sometimes drastically, sometimes more subtly. It is
impossible to ignore the complexity of life and represent a multi-dimensional phenomena as a rolling
dot model. |
The Palaeolithic Continuity Theory: The Linguistic View
Writing a half century after Gimbutas proposed the Kurgan theory, Alinei (2001)
asserts that a great IE invasion in the Chalcolithic, triggering a total ethnic and linguistic
substitution on a continental scale, is simply inconceivable. He suggests that the greater part of
the common Neolithic IE lexicon, (i.e. loan words designating innovative devices like the plow, the
yoke, the wheel, some domesticated animals, plants, and some metals), was already diversified in
almost all the IE languages during the Mesolithic and the Neolithic.
With respect to the Anatolian hypothesis, Alinei (2001) observes that limited
migrations in the Balkans and Central Europe from Anatolia, even over a few millennia, are not
sufficient explanation for the development and differentiation of IE languages in Europe.
Moreover, these migrations cannot explain the relatively large number of NIE toponyms in the Aegean area and the NIE words in Greek and other languages in southern Italy, Sicily, Sardinia,
Corsica, and Spain. These observations, according to Alinei, support the hypothesis that the
populations coming from Anatolia were NIE speakers and that the Neolithic in Europe was a period of
complex acculturation and geographical differentiation in which small migrating groups played a
limited role with respect to the populations already inhabiting Europe.
| It became a continent-wide sport to fancy an IE etymology to every word encountered in the
would-be IE languages, and the NIE inventory grows smaller, while the IE stock grows wider and
vaguer. The etymological ingenuity makes great strides in reducing the relatively large number of NIE
words to relatively insignificant number, inadvertently pulling out the NIE carpet from the PCT
theory. Patriotism and enthusiasm can turn mountains upside down. Since it was positively proved
that the world was created 7,000 years ago, and the IE theories already extended IE to 11,000 ybp,
we have already within a range that allows to stipulate that the Snake, Adam, and Eve probably
conversed in IE to resolve a problem of biting apples. |
The fact that the greater portion of the grammatical structure of Celtic,
Germanic, Italic, Greek, Illyrian, and Balto-Slavic is different from IE grammatical structures
indicates that they could not have been formed in the Chalcolithic or Eneolithic. Alinei (2001)
indicates that that the only possible solution to the linguistic conundrum is offered by the
Palaeolithic Continuity Theory (PCT). PCT is supported by paleoanthropologists who have concluded
that that not only Homo erectus
but also Homo habilis and perhaps even
Australopitecus
were able to speak (Tobias, 1996). Some researchers in the cognitive sciences have reached the
same conclusion, (i.e., to explain the innate character of human language it is necessary that
Australopithecus
had some capacity for language [Pinker, 1994]). Thus, the structural portions of all human
languages, including PIE (i.e. words, affixes, syntax) allegedly were formed a long time ago in
Africa as part of human evolution.
Alinei (2001) believes that Neolithic Europe would have been occupied by IE, NIE
and Uralic peoples, though the NIE speakers would have influenced the IE languages only by contact
and adstrates. Although he excludes the possibility of a massive invasion of Europe during the
Chalcolithic or Neolithic, Alinei (2001) notes that an important hybridization took place in
southern Europe at the beginning of the Neolithic as a result of the infiltration of NIE populations
and the migrations of the Kurgan peoples during the Chalcolithic. Other hybridizations took place
during the Bronze Age. However, these hybridizations would have altered the languages and cultures
of the IE populations only superstratically.
Alinei (2001) asserts that Celtic and north Celtic peoples occupied western
Europe, including Brittany and Ireland, before the retreat of the glaciers, and that they created
megalithism and the TBK cultures. During the Palaeolithic, the Italide or Italoide ethnolinguistic peoples occupied southern Europe from the Iberian Peninsula to
Dalmatia. During the Neolithic, the Balkan area was influenced by NIE migrant groups of farmers, who
created the Balkan Sprachbund, (i.e. the Balkan group of languages: Greek,
Serbian, Bulgarian, Macedonian, Albanese, and Romanian).
107
According to Alineli (2001) the Kurgan culture introduced Turkic, not Iranian,
influences to IE languages, and the border between the Trypillia and the Srednyj Strog cultures is
the border between Slavic and Turkic cultures. In this theory, the late Combat Axe population were
the IE peoples influenced by the Kurgan culture. Furthermore, according to the Uniformity Principle of historical linguistics, the languages of Europe during the Bronze Age correspond to the
languages of modern Europe (i.e., the areas of Bronze Age civilizations correspond to dialectal
language areas, which in turn correspond to each IE language).
The Palaeolithic Continuity Theory: The View of DNA Genealogy
The Palaeolithic Continuity Theory appears incompatible with the history of
Europe based on DNA genealogy data.
The PCT places the origin of PIE languages in Europe in the Upper Palaeolithic
(minimum 10,000 ybp), and links it to the arrival of people in Europe from Africa; it proposes the
continuity of peoples and languages in Europe for the last at least 10,000 ybp. This view is
contradicted by DNA genealogy data.
The only parts of the PCT which find support from DNA genealogy are:
— PIE languages arrived in Europe around 10,000 ybp; they did not, however, arrive
from Africa, but from Asia, via Anatolia.
— Words designating innovative devices, domesticated animals, plants and some
metals, were already diversified in IE languages, and were not brought by R1b "invaders" who arrived in Europe at the beginning of the 5th millennium bp.
| In the context of the IE languages, “diversified” can't mean dialectal diversification that
creates phonetical versions of the common word, it only can mean that different IE languages had
different words for the same innovative devices, domesticated animals, plants and metals. In each
case, the source of the diversification for the 5th millennium bp can't be established due to
the absence of any recorded data for the majority of the IE languages in the next 3 millennia, and
that denies an opportunity to establish if the same diversification existed in the 5th and 1st
millennium bp. The same R1b word could be brought in 5th and 1st millenniums bp, and be semantically
and phonetically exactly the same. This is a grey area that allows unlimited speculations and
beliefs. |
According to DNA genealogy data, genealogical lineages or haplogroups, and
languages in Europe have not shown a continuous pattern. In fact, according to DNA genealogy data:
— IE (R1a) populations fled from Europe to the Russian Plain around 4600 ybp.
There were at least ten R1a tribes each with a distinct subclade/SNP and/or branch of haplotypes,
which migrated back to Europe after 3000 ybp.
— Haplogroup G populations were almost completely eliminated in Europe between
4500 and 4000 ybp, apparently by the arrival of the Arbins (R1b); the survivors fled to Asia Minor,
Mesopotamia and the Caucasus. There are excavated haplotypes of E-V13 dated 7000 ybp; however, the
common ancestor of contemporary E-V13 bearers lived only 3500 ybp, indicating a population bottleneck.
— Haplogroup I1 populations were almost completely eliminated in Europe between
4500 and 4000 ybp; they went through a severe population bottleneck until around 3600 ybp, which was
a new beginning for I1 haplotypes in Europe.
— Haplogroup I2 populations were almost completely exterminated in Europe 4500
ybp, and the survivors fled to England and Ireland, and to eastern Europe. Present day I2
populations have common ancestors at 4800 ybp and 2300 ybp, respectively (Klyosov, 2012c).
— NIE speakers (R1b) arrived in Europe near the Pyrenees around 4800 ybp; they
arrived at the Apennines and the Balkans from the Pontic steppes, around 4500 ybp. These migrations caused major disruptions in the populations and languages of Old Europe.
There are other aspects of PCT that are questionable in the light of DNA
genealogy:
— The notion that a few millennia in the Neolithic and a limited number of
migrations to the Balkans and central Europe from Anatolia were not sufficient for the development
and differentiation of the IE languages in Europe is questionable. In fact, IE speakers (R1a) were
in Europe from about 10,000 to about 4500 ybp. Thus, it cannot be assumed that there was a too short
time for "development" and "differentiation" of IE languages.
— The proposal of the PCT that the arrival of IE languages from Anatolia cannot
explain the relatively large number of NIE toponyms in the Aegean area and the NIE words in Greek
and other languages of South Italy, Sicily, Sardinia, Corsica and Spain (see above; Alinei, 2001) is
questionable. The IE speakers (R1a) did not come to an empty Europe, there already were NIE
populations of haplogroups E, F, G, I1, I2, J2, K, etc. So, IE languages were very likely introduced
in a NIE speaking Europe. Moreover, the PCT assumption that the populations coming from Anatolia
were NIEs (see above) is contradicted by DNA genealogy data. It might well be, though, that some
other haplogroups/tribes speaking NIE languages also migrated to Europe about between 10,000 and
9000 ybp; nevertheless, even if that had happened, it would not change the language landscape of
ancient Europe.
— The PCT suggestion that the structural portions of all human languages, formed
long ago in Africa in connection with human evolution (see above) appears erroneous. Nobody can
responsibly exclude the idea that H. habilis and
Australopithecus were able to speak; however, DNA genealogy has shown that
non-Africans do not have "African" SNPs on their Y chromosomes (Klyosov & Rozhanskii, 2012; Klyosov
et al., 2012). Africans and non-Africans have plenty of SNP-mutations from a common ancestor of
humans and chimpanzees; however, non-Africans have apparently, not received them from the Africans
(ibid.). As a consequence of this lack of DNA data, it is hard to imagine that African languages
could have evolved into PIE languages. Overall, it is highly ques-tionable that "anatomically modern
homo" arose in Africa (Klyosov & Rozhankii, 2012; Klyosov et al., 2012; Bednarik, 2012, 2013), see
also
Figure 1 1.
— The suggestion of the PCT that Celtic and north Celtic populations occupied
Western Europe, including Brittany and Ireland, as long ago as before the retreat of glaciers
appears erroneous. According to DNA genealogy data, Celtic IE languages reached England and
Ireland in the 3rd millennium bp. Their languages were imposed on the existing NIEs (R1b). This
explains why Indo European languages are spoken today in Britain and Ireland by R1b populations
(around 90% and above of today populations) plus a few (singular per cent) of R1a, I1, I2, and other
minor haplogroups populations in Britain and Ireland.
Earlier Genetic Studies
This section describes a number of erroneous statements made in the early stages
of genetic genealogy (also called genogeography and/or population genetics), in the 1990s and 2000s.
Some of these statements still carry weight in linguistics.
108
The founding fathers of genetic genealogy claim, for example, that bearers of R1b
lived in Europe 30,000 ybp (Wells et al., 2001; Wells, 2006), or between 40,000 and 35,000 ybp years
ago (Semino et al., 2000). The main reason for this presumption is, apparently, that if R1b
populations live in Europe now, they have lived there always. The claim that there were R1b tribes
in Europe about 30,000 ybp, stuck for 15 years and even continues to be cited in contemporary
population genetics literature. The date has been cross-cited hundreds of times in academic publications. However, according to DNA genealogy, R1b tribes arrived to Europe only between 4800 and
4500 ybp (Klyosov, 2012b).
The founding fathers of genetic genealogy claimed initially that haplogroup R1a
arose in the southern Russian steppes about 15,000 ybp (Wells et al., 2001). Five years later, the
estimate date was changed (without explanation) to 10,000 ybp (Wells, 2006). In fact, both dates
were invented. Without offering any substantiation for the claim, the founders postulated that the
oldest R1a bearers survived the Ice Age in a Ukrainian refuge (Semino et al., 2000). As a result,
R1a was called the Ukrainian haplogroup (e.g., Wiik, 2008) for years — without any justification.
Genetic genealogists claim, without any supporting facts (for a mini-review see
Klyosov et al., 2012) that genetic data show that man left Africa some 70,000 ybp (or 50,000 or
60,000 ybp). They make no calculations based on Y chromosome markers. They base their "Out of
Africa" theory on the comparative diversity of African haplogroups. However, diversity as a crite
rion of age is valid only in closed systems. In open systems, such as Africa in this particular
case, diversity is a consequence of the mixing of bearers of different Y chromosomes. Unfortu
nately, these erroneous dates have been used in academic literature from the 1990s to the present
time. More recent genome studies have shown that there is a wide gap between the African genome
exemplified with indigenous hunter-gatherer peoples (Schustler et al., 2010; Lachance et al., 2012,
and ref. therein; Klyosov et al., 2012), and the non-African genome, as, in fact, should follow from
Figure 1
above. There are no indications that non-Africans descended from Africans. African SNPs are absent,
for example, in Europeans (Klyosov & Rozhankii, 2012; Klyosov et al., 2012). Klyosov et al. (2012)
have shown that the stream of SNP mutations from a common ancestor with chimpanzees goes to the
α-haplogroup, from which the African lineage (haplogroup A) split around 160,000 ybp,
and evolved in a separate Y-chromosomal lineage from the Europeoid lineage. Another archaic
African lineage split even earlier, some 200,000 ybp or perhaps some 350,000 ybp (Mendez et al.,
2013); bearers of this archaic lineage still live in Africa. In other words, the "Out of Africa"
hypothesis has presented a distorted pattern not only of the origin of man but also of the de
velopment of human languages (Klyosov & Rozhanskii, 2012; Klyosov et al., 2012; Bednarik, 2012,
2013).
The population geneticists of the 1990s-2000s, have tried, apparently, to match
the historical convictions of those decades by bending their DNA-based theories. They uncritically
consider gradients of frequency (or clines), which can always be found for whatever reason,
including population bottlenecks, ignoring the existence of downstream subclades. In many studies
(Hammer, 2009; Underhill, 2009; Zhivotovsky, 2004) erroneous mutation rates were employed (e.g.,
population rate constants or Zhivotovsky mutation rates — which increase the actual number of
years to common ancestors by 300% - 400%). As a result, the dating of populations are inflated by a
factor of 3 or 4. Using these measures, Indo Europeans first appeared in India 14,000 ybp rather
than 3500 ybp. There are dozens of examples of this kind in the literature.
Similarly, Semino et al. (2000) concluded that some European peoples (e.g., the
Basques) are genetically different from others. But the majority of contemporary Basques belong to
haplogroup R1b and share with about 60% of all Europeans the same arrival-in Europe-date. This
conclusion was recently confirmed using a genome-wide study of the Basques, according to which
Basques are not genetic outlier among European populations (Laayouni et al., 2010). In fact,
southern Europe has many Palaeolithic haplogroups, such as E, F, G, K, J, which passed through a
severe population bottleneck around 4500 ybp apparently as a result of the Arbins’ (R1b) arrival
in Europe. Similarly, northern Europe has Palaeolithic haplogroups, such as I1, which passed the
same bottleneck and started to recover only about 3600 ybp. We do not know their "genetic components" before that.
Conclusion
To sum up, early genetic studies of the origin of Europeans often present
superficial conclusions based on scarce data that has not been subjected to serious scientific
scrutiny. DNA genealogy has not only enabled us to re-construct migration and settlement patterns
in ancient Europe, it has also permitted us to put the leading linguistic theories under scrutiny.
We have been able to disprove the Kurgan theory and the Palaeolithic Continuity Theory and bring
into question the "Out of Africa" hypothesis. We have also been able to fine tune the Vasconic and
Anatolian theories.
Acknowledgements
The authors are indebted to Dr. Judith Remy Leder for her valuable help with the
preparation of the manuscript.
REFERENCES
Alinei, M. (2001). An alternative model of the people-related sources in European
languages: The "continuity theory". In B. Mondadori (Ed.), The first roots of Europe (pp. 177-234). (in Italian)
Anikovich, M. V., Sinitsyn, A. A., Hoffecker, J. F., Holliday, V. T., Popov, V.
V., Lisitsyn, S. N., Forman, S. L. et al. (2007). Early upper paleolithic in Eastern Europe and
implications for the dispersal of modernhumans.
Science, 315,
223-226. doi:10.1126/science.1133376
Baldi, P., & Page, R. B. (2006). Review europa vasconica — Europa semitica.
Lingua, 116,
2183-220. (in German)
Bednarik, R. G. (2012). The origins of human modernity.
Humanities, 1,
1-53.
Bednarik, R. G. (2013). Pleistocene palaeoart of Africa. Arts, 2, 6-34.
Benazzi, S., Douka, L., Fornai, C., Bauer, C. C., Kullmer, O., Svoboda, J., Pap,
I., et al. (2011). Early dispersal of modern humans in Europe and implications for Neanerthal
behaviour. Nature, 479,
525-528. doi:10.1038/nature10617
Bököny, S. (1997). Horses and sheep in East Europe. In S. N. Skomal, & E. C.
Polomé (Eds.), Proto-Indo-European: The archaeology of a linguistic problem. Studies in
honour of Marija Gimbutas
(pp. 136 - 144). Washington DC: Institute for the Study of Man.
Boric, D., & Price, T. D. (2013). Strontium isotopes document greater human
mobility at the start of the Balkan Neolithic.
Proceedings of the National Academy of Sciences of the USA, 110, 3298-3303.
doi:10.1073/pnas.1211474110
Bouckaert, R. et al. (2012). Mapping the origins and expansion of the
Indo-European language family.
Science, 337,
957-960. doi:10.1126/science.1219669
109
Schustler, S. C., Miller, W., Ratan, A., Tomsho, L. P., Giardine, B., Kasson, L.
R., Harris, R. S. et al. (2010). Completer Khoisan and Bantu genomes from southern Africa.
Nature, 463,
943-947. doi:10.1038/nature08795
Diakonov, J. M. (1984). On the original home of the speakers of Indo European.
Soviet Anthropology and Archaeology, 23, 5-87.
doi:10.2753/AAE1061-195923025
Dolgopolsky, A. B. (1987). The Indo-European homeland problem.
Mediterranean Archaeological Review, 3,
7-31.
Dolgopolsky, A. B. (1993). More about the Indo-European homeland problem.
Mediterranean Archaeological Review, 6-7, 251-272.
Gamkrelidze, T. V., & Ivanov, V. V. (1984).
Indo-European and the Indo-Europeans. Tbilisi: Tbilisi State University.
Gamkrelidze, T. V., & Ivanov, V. V. (1995). Indo-Europeans and the
Indo-Europeans.. In J. Nichols, & M. de Gruyter (Eds.),
A reconstruction and historical analysis of a proto-language and a protoculture
(1128 pp). Berlin-New York.
Gimbutas, M. (1991).
The civilization of the goddess: The world of Old Europe, Harper. 544 pp.
Gimbutas, M. (1956). The prehistory of Eastern Europe. Mesolithic, neolithic and
copper age cultures in Russia and the Baltic area. American school of prehistoric researches.
Harvard University Bulletin, 20,
Cambridge: Peabody Museum,
Gimbutas, M. (1994).
Das ende alteuropas (The end of the Old Europe) (135 pp). Budapest: Verlag des
Instituts fur Sprachwissensvhaft der Universitat Innsbruck. (in German)
Gimbutas, M. (1997). The fall and transformation of Old Europe: Recapitulation.
Journal of Indo-European Studies, 18, 351-372.
Gray, R. D. & Atkinson, Q. D. (2003). Language-tree divergence times support the
Anatolian theory of Indo-European origin.
Nature, 426, 435-439. doi:10.1038/nature02029
Gray, R. D., Atkinson, Q. D., & Greenhill, S. J. (2011). Language evolution and
human history: What a difference a date makes. Philosophical Transactions of the Royal
Society B, 366, 1090-1100.
doi:10.1098/rstb.2010.0378
Hammer, M. F., Behar, D. M., Karafet, T. M., Mendez, F. L., Hallmark, B., Erez,
T., Zhivotovsky, L. A. et al. (2009). Extended Y chromosome haplotypes resolve multiple and unique
lineages of the Jewish priesthood.
Human Genetics, 126, 707-717.
doi:10.1007/s00439-009-0727-5
Higham, T., Compton, T., Stringer, C., Jacobi, R., Shapiro, B., Trinkaus, E.,
Chandler, B. et al. (2011). The earliest evidence for anatomically modern humans in northwestern
Europe. Nature, 479,
521-524. doi:10.1038/nature10484
Keyser, C., Bouakaze, C., Crubezy, E., Nikolaev, V. G., Montagnon, D., Reis, T.,
& Ludes, B. (2009). Ancient DNA provides new insights into the history of south Siberian Kurgan
people. Human Genetics, 126, 395-410. doi:10.1007/s00439-009-0683-0
Klyosov, A. A. (2009). DNA genealogy, mutation rates, and some historical
evidences written in Y-chromosome. II. Walking the map.
Journal of Genetic Genealogy, 5, 217-256.
Klyosov, A. A. (2010). Haplogroup I. Proceeding of Academy of DNA Genealogy, 3, 96-158.
Klyosov, A. A. (2012a). Microsatellites and genes of Y-chromosome.
Proceeding of Academy of DNA Genealogy, 5, 911-913.
Klyosov, A. A. (2012b). Ancient history of the Arbins, bearers of haplogroup
R1b, from Central Asia to Europe, 16,000 to 1500 years before present.
Advances in Anthropology, 2,
87-105. doi:10.4236/aa.2012.22010
Klyosov, A. A. (2012c). Dinaric (East-European) and the "Isle" branches of
haplogroup I2a. Proceeding of Academy of DNA Genealogy, 5, 1304-1317.
Klyosov, A. A., & Rozhanskii, I. L. (2012a). Re-examining the "Out of Africa"
theory and the origin of Europeoids (Caucasoids) in light of DNA genealogy.
Advances in Anthropology, 2, 80-86.
doi:10.4236/aa.2012.22009
Klyosov, A. A., & Rozhanskii, I. L. (2012b). Haplogroup R1a as the proto
Indo-Europeans and the legendary Aryans as witnessed by the DNA of their current descendants.
Advances in Anthropology, 2, 1 - 13. doi:10.4236/aa.2012.21001
Klyosov, A. A., Rozhanskii, I. L., & Ryanbchenko, L. E. (2012). Reexamining the
Out-of-Africa theory and the origin of Europeoids (Caucasoids). Part 2. SNPs, haplogroups and
haplotypes in the Y chromosome of Chimpanzee and Humans.
Advances in Anthropology, 2,
198-213. doi:10.4236/aa.2012.24022
Klyosov, A. A. (2013). Subclade R1a-L342-L657 beyond Ancient Urals.
Proceeding of Academy of DNA Genealogy, 6,
446-451.
Krahe, H. (1954).
Language and prehistory: European history according to the testimony of the language.
Heidelberg: Quelle und Meyer. (in German)
Krahe, H. (1964).
Our oldest river names. Wiesbaden: Harrassowitz.
Kuzmenko, Y. (2011).
Earlier Germans and their neighbours: Linguistics, archaeology, genetics. St.
Petersburg: Nestor-History.
Laayouni, H., Calafell, F., & Bertranpetit, J. (2010). A genome-wide survey does
not show the genetic distinctiveness of Basques.
Human Genetics, 127,
455-458. doi:10.1007/s00439-010-0798-3
Lacan, M., Keyser, C., Ricaut, F.-X., Brucato, N., Tarrús, J., Bosch, A.,
Guilaine, J. et al. (2011). Ancient DNA suggests the leading role played by men in the Neolithic
dissemination. Proceedings of the National Academy of Sciences of the United States of
America, 108,
18255-18259. doi:10.1073/pnas.1113061108
Lachance, J., Vernot, B., Elbers, C. C., Ferwerda, B., Froment, A., Bodo, J.-M.,
Lema, G. et al. (2012). Evolutionary history and adaptation from high-coverage whole-genome
sequences of diverse African hunter-gatherers.
Cell, 150, 1-13.
doi:10.1016/j.cell.2012.07.009
Li, C., Li, H., Cui, Y., Xie, C., Cai, D., Li, W., Mair, V. H. et al. (2010).
Evidence that a west-east admixed population lived in the Tarim Basin as early as the early Bronze
Age. BMC Biology, 8.
doi:10.1186/1741-7007-8-15
Marler, J. (2001). L’Eredità di Marija Gimbutas: Una ricerca archeomitologica
sulle radici della civiltà europea. In B. Mondadori (Ed.),
The first roots of Europe
(pp. 3-22). (in Italian)
Mendez, F. L., Krahn, T., Schrack, B., Krahn, A.-M., Veeramah, K. R., Woerner, A.
E., Fomine, F. L. M. et al. (2013). An African American paternal lineage adds an extremely ancient
root to the human Y chromosome phylogenetic tree.
The American Journal of Human Genetics, 92,
454-459.
Pinker, S. (1994).
The language instinct: The new science of language and mind. London-New York: Penguin
Books.
Prat, S., Pean, S. C., Crepin, L., Drucker, D. G., Puaud, S. J., Valladas, H.,
Galetova-Laznickova, M. et al. (2011). The oldest anatomically modern humans from far southeast
Europe: Direct dating, culture and behavior.
PLoS ONE, 6,
e20834. doi:10.1371/journal.pone.0020834
Pringle, H. (2012). News & analysis: New method puts elusive Indo-European
homeland in Anatolia. Science, 337,
902.
doi:10.1126/science.337.6097.902
Renfrew, C. (2001). Origini indoeuropee: Verso una sintesi. In B. Mondadori
(Ed.), The first roots of Europe. (in Italian)
Rozhanskii, I. L., & Klyosov, A, A. (2012). Haplogroup R1a, its subclades and
branches in Europe during the last 9000 years.
Advances in Anthropology, 2,
139-156. doi:10.4236/aa.2012.23017
Ryan, W., & Pittman, W. (1998).
Noah’s flood: The new scientific discoveries about the event that changed history. New
York: Simon & Schuster.
Ryder, R. J., & Nicholls, G. K. (2011). Missing data in a stochastic Dollo model
for binary trait data and its application to the dating of Proto-Indo-European.
Journal of the Royal Statistical Society: Series C (Applied Statistics), 60, 71-92.
doi:10.1111/j.1467-9876.2010.00743.x
Schmid, W. P. (1987). "Indo-European" — "Old European". In S. N. Skomal., & E. C.
Polomé (Eds.), Proto-Indo-European: The archaeology of a linguistic problem: Studies in
honour of Marija Gimbutas
(pp. 322-338). Washington: Institute of the Study of Man.
Schmid, W. P. (2001). Was Gewässernamen in Europa besagen.
Akademie-Journal, 2,
42-45.
Semino, O., Passarino, G., Oefner, P. J., Lin, A. A., Arbuzova, S., Beckman, L.
E., De Benedictis, G. et al. (2000). The genetic legacy of Paleolithic Homo sapiens sapiens in
extant Europeans: A Y chromosome perspective.
Science, 290,
1155-1159.
Sharma, S., Rai, E., Sharma, P., Jena, M., Singh, S., Darvishi, K., Bhat,
A. K. et al. (2009). The Indian origin of paternal haplogroup R1a1* substantiates
the autochthonous origin of Brahmins and the caste system.
Journal of Human Genetics, 54,
47-55.
110
Sturtevant, F. H. (1962). The Indo-Hittite hypothesis.
Language, 38,
376-382. doi:10.2307/410871Tobias, P. V. (1996). The evolution of the brain, language and cognition. In F.
Facchini (Ed.), Colloquium VIII: Lithic-industries, language and social behaviour in the
first human form
(pp. 87-94), Forli.
Tomezzoli, G. T., & Kreutz, J. (2011). The linguistic position of the Tocharian.
Proceedings of the 9th International Topical Conference: Origin of Europeans,
Ljubljana, 67-86.
Trask, R. L. (1995). Origin and relatives of the Basque language: Review of the
evidence. In: J. L. Hualde et al. (Eds.),
Toward a history of the Basque language
(pp. 65-77). Amsterdam: Benjamin.
Trask, R. L. (1996).
The history of basque
(480 pp). London, New York: Routledge.
Underhill, P. A., Myres, N. M., Rootsi, S., Metspalu, M., Zhivotovsky, L. A.,
King, R. J. et al. (2009). Separating the post-glacial coancestry of European and Asian Y
chromosomes within haplogroup R1a.
European Journal of Human Genetics, 18, 479-484.
doi:10.1038/ejhg.2009.194
Vennemann, T. (2003).
Europa Vasconica-Europa semitica
(999 pp). Berlin: Mouton de Gruyter.
Wells, R. S., Yuldasheva, N., Ruzibakiev, R., Underhill, P. A., Evseeva, I.,
Blue-Smith, L., Jin, L. et al. (2001). The Eurasian heartland: A continental perspective on
Y-chromosome diversity. Proceedings of the National Academy of Sciences of the United
Stated of America, 98,
10244-10249. doi:10.1073/pnas.171305098
Wells, S. (2006). Deep ancestry: Inside the genographic project.
National Geographic, 248,
247 pp.
Wiik, K. (2008). Where did European men come from?
Journal of Genetic Genealogy, 4,
35-85.
Zhivotovsky, L. A., Underhill, P. A., Cinnoglu, C., Kayser, M., Morar, B.,
Kivisild, T., Scozzari, R. et al. (2004). The effective mutation rate at Y chromosome short tandem
repeats, with application to human population-divergence time.
The American Journal of Human Genetics, 74, 50-61.doi:10.1086/380911
Zimmer, H. (1898). Matriarchy among Picts. In G. Henderson, & G. Zimmer (Eds.),
Leabhar nan gleann (The book of the glens)
(pp. 1-42). Edinburgh: Norman Macleod
111
|


{kind=link}