Miroslava V. Derenko et. al.
Presence of Mitochondrial Haplogroup X in Altaians from South Siberia
Miroslava V. Derenko, Tomasz Grzybowski, Boris A. Malyarchuk, Jakub Czarny, Danuta Miscicka-Sliwka, Ilia A. Zakharov
Am. J. Hum. Genet. 69:237-241,
2001 Letters to the Editor
© 2001 by The American Society of Human Genetics. All rights reserved
Abstracts related to the Türkic History
Links
Foreword
Miroslava V. Derenko et. al.
Presence of Mitochondrial Haplogroup X in Altaians from South Siberia
237
To the Editor:
For American Indians, extensive RFLP and HVSI sequence analysis has unambiguously identified four major founding mtDNA haplogroups (A, B, C, D), which account together for ~97% of modern American Indian mtDNAs (Wallace 1995). Examination of the distribution of the four founding lineage haplotypes (A, B, C, and D) in American Indian populations (both contemporary and ancient) shows that all four lineages were present in the New World prior to European contact (Wallace 1995; Lalueza et al. 1997; Stone and Stoneking 1998), thus indicating that all American Indian mtDNAs are apparently descended from these four founding lineages. mtDNAs apparently not from haplogroups A-D may result from recent admixture with non-American Indians or may represent additional American Indian founding mtDNA lineages.
A striking example of the presence in American Indians of genotypes not from haplogroups A-D is
haplogroup X. This haplogroup represents a minor founding lineage that is restricted in
distribution to northern Amerindian groups, including the Ojibwa, the Nuu-Chah-Nulth, the Sioux,
and the Yakima, as well as the Na Dene-speaking Navajo (Brown et al. 1998). Unlike haplogroups
A-D, haplogroup X is also found at low frequencies of ~4% in western Eurasian populations.
Despite a shared consensus RFLP haplotype, substantial genetic differences exist between the
American Indian and European haplogroup X mtDNAs. Phylogenetic analysis and coalescence estimates
for American Indian and European haplogroup X mtDNAs exclude the possibility that the occurrence
of haplogroup X in American Indians is due to recent European admixture. They also clearly indicate
that the two branches/subgroups are distantly related to each other and that considerable genetic
substructure exists within both groups (Brown et al. 1998).
238
| Sample | Origin | RFLP Haplotype a 111 1046 7345 1961 5457 ccse |
HVSI, HVSII Sequence b 666666 11112201122333 88892775916001 23933833553995 a aba |
| CRS | +–--AAT- | AAT-C C AAT AA | |
| ALT16 | Northern Altaians | CCCCTTGGCGGCCC | |
| ALT43 | Northern Altaians | CCC.TTGGC.G..C | |
| ALT81 | Southern Altaians | CCCCTTGGC.GCCC | |
| ALT161 | Southern Altaians | CCC.TTGGC.GCCC | |
| ALT171 | Southern Altaians | CCC.TTGGC.GCCC | |
| ALT188 | Southern Altaians | CCC.TTGGC.GCCC | |
| ALT208 | Southern Altaians | CCC.TTGGC.GCCC |
a Restriction-endonuclease sites are indicated as
follows: c p DdeI, e p HaeIII, s p AccI. A dot (.) denotes identity with the Cambridge reference
sequence (Anderson et al. 1981); a plus sign ( + ), which denotes a site gain, or a minus sign ( — ),
which denotes a site loss, indicates deviation from the CRS.
b HVSI (nucleotide positions 15991-16400) and HVSII (nucleotide positions 20-420) were sequenced.
A dot (.) indicates identity with the CRS, and a dash ( — ) indicates nucleotide insertion.
Haplogroup X is remarkable in that it has not been found in Asians, including Siberians, suggesting that it may have come to the Americas via a Eurasian migration. The virtual absence of haplogroup X in eastern and northern Asia raises the possibility that some American Indian founders were of European ancestry. In that case, as it has been proposed, haplogroup X was brought to America by the eastward migration of an ancestral white population, of which no trace has so far been found in the mtDNA gene pool of modern Siberian/eastern Asian populations (Brown et al. 1998).
However, it should be stressed that mtDNA-variability studies of the populations living in this major geographic area were performed on a limited number of populations. Some regions remain poorly sampled, and more extensive sampling is required. Moreover, some key markers, including those defining the X-haplogroup sequences, have not been typed for many different populations. These limitations do not allow correct definition of the phylogenetic status of mtDNA lineages.
To extend the survey of Asian mtDNAs for the presence of haplogroup X, we screened the mtDNAs of a total of 790 individuals for the RFLP markers (-1715 DdeI, -10394 DdeI, +14465 AccI, and +16517 HaeIII) that define this lineage. These individuals comprised 10 aboriginal Siberian populations: Buryats (n = 105) , Tuvinians (n = 111), Koryaks (n = 35), Evens (n = 65), Yakuts (n = 62), Khakassians (n = 54), Shors (n = 42), Sojots (n = 34), Altaians (n = 202), and Evenks (n = 80). All individuals belonged to the indigenous population of the regions studied, were unrelated, and stated that their maternal grandmother had been born in the area considered for this study.
Haplogroup X mtDNAs were detected only in Altaians, at a frequency of 3.5%.
The haplogroup X status of these haplotypes was confirmed through HVSI and HVSII mtDNA sequencing (table 1). All Altaian X mtDNAs harbored the consensus haplogroup X motif: -1715 DdeI, +14465 AccI, +16517 HaeIII, 16189C, 16223T, 16278T, 73G, 153G, 195C, 263G, relative to the Cambridge reference sequence (Anderson et al. 1981) and differed from each other by length-polymorphism mutations at nucleotide positions 16193, 309, and 315. One of these X mtDNAs (ALT16) also harbored a 215G variant (Table 1) that has not been observed in either American Indian or European X haplotypes. It should also be noted that none of the Altaian X mtDNAs harbored the 225A variant, which is a marker for a major part of haplogroup X (Brown et al. 1998).
Analysis of published data on European HVSI and HVSII mtDNA sequences (Piercy et al. 1993; Calafell et al. 1996; Torroni et al. 1996; Brown et al. 1998; Lutz et al. 1998; Parson et al. 1998; Rousselet and Mangin 1998; Helgason et al. 2000) demonstrates that the overwhelming majority of X haplotypes (23 of 25 X sequences) harbor the 225A variant. In contrast, the X haplotypes without 225A have been observed mostly in American Indians (11 of 14 Ojibwa; see Table 1 in Brown et al. 1998). Nevertheless, the X mtDNAs that we detected in the Altaian sample do not bear the 16213A and 200G variants that are characteristic of most American Indian haplogroup X mtDNAs (Brown et al. 1998).
Figure 1 illustrates the reduced median network, constructed by means of the median algorithm of Bandelt et al. (1995), encompassing the HVSI and HVSII variation observed in the American Indian, European, and Altaian haplogroup X mtDNAs. The network suggests that European and American Indian haplogroup X mtDNAs are separated into two major branches, whereas the majority of Altaian X mtDNAs appear to be very similar to the root of haplogroup X phylogeny, differing from it by one step (loss of 225A). The network further suggests that the Altaian X haplotypes occupy the intermediate position between European and American Indian haplogroup X mtDNA lineages (Fig. 1).
The Altaians, the native people of Altai Republic (south Siberia) number up to 60,000 persons. "Altaians" is the common denomination for seven formerly distinct Turkic-speaking groups: the Altai-Kiji, Teleuts, and Telengits, who are southern Altaians, and the Chelkans, Kumandins, Tubalars, and Maimalars, who are northern Altaians. The differences between southern and northern Altaians are well established, on the basis of anthropological, linguistic, and classical genetic-marker studies (Potapov 1969; Alexeev and Gohman 1984; Luzina 1987). Anthropologically, southern Altaians are typical central Asian Mongoloids (like Mongolians, Yakuts, and Buryats), whereas northern Altaians exhibit some Caucasoid anthropological features, similar to those of other Türkic, Ugric and Samoyedic groups.
Figure 1 Reduced median network of haplogroup X mtDNAs, as defined by HVSI and HVSII variation.
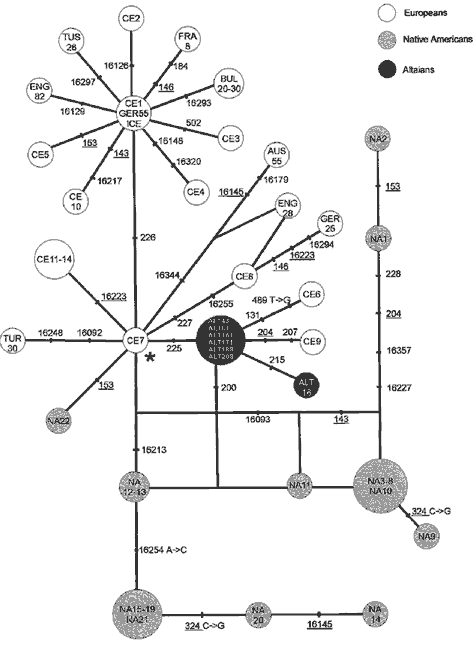 |
RFLP status of the
sequences is known for American Indians (NA; Brown et al. 1998); |
240
The Altai region was populated during the Lower Paleolithic, and there is ample evidence of settlement during the Middle Paleolithic. It was proposed by anthropologists that, at least from the Neolithic, the territories of Altai and Sayan region were populated by mixed tribes with Caucasoid and Mongoloid anthropological features, but later they were replaced by Mongoloid populations of central Asian origin (Alexeev and Gohman 1984). The analysis of the tribal structure of Southern Altaians has shown that the present-day Altaians have retained their native language and ethnic identity.
They have begun to mix with other ethnic groups (mostly Russians and Kazakhs) only recently, so the interethnic admixture is estimated to be 15% (Luzina 1987; Osipova et al. 1997). The haplogroup X mtDNAs have not been found in populations of central Asia, including Kazakhs, Uigurs, and Kirgizs (Comas et al. 1998).
Since the frequency of haplogroup X in Russians is extremely low (3 of 336; Orekhov et al. 1999; Malyarchuk and Derenko 2000; authors' unpublished data), the recent European admixture cannot explain the presence of haplogroup X in the Altaians. Hence, the results of the present study allow us to suggest that haplogroup X was the part of the ancestral gene pool for Altaian populations, being found both in northern and southern Altaians.
Recently, the mtDNA studies have shown that both northern and southern Altaians exhibit all four Asian and American Indian-specific haplogroups (A-D) with frequencies of 57.2% (Sukernik et al. 1996) and 46.8% (Derenko et al. 2000a), respectively, exceeding those reported previously for Mongolians, Chinese, and Tibetans. Therefore, they may represent the populations which are most closely related to New World indigenous groups. Since the detection of all four haplogroups (A-D) in an Asian population is thought to be a first criterion in the identification of a possible New World founder, the candidate source population for American Indian mtDNA haplotypes therefore may include the populations originating in the regions to the southwest and southeast of Lake Baikal, including the Altai Mountain region (Derenko et al. 2000b). The presence of X mtDNAs in Altaians is generally consonant with the latter conclusion.
Because the location and identification of the population that was the source of the founding lineages for the New World is a question of considerable interest, several studies on Y-chromosomal DNA polymorphism were performed recently to investigate Pleistocene male migrations to the American continent (Underhill et al. 1996; Lell et al. 1997; Karafet et al. 1999; Santos et al. 1999).
It has been shown that the major Y haplotype present in most American Indians could be traced back to recent ancestors they have in common with Siberians: namely, the Kets and Altaians, from the Yenisey River Basin and the Altai Mountains, respectively (Santos et al. 1999). Similarly, based on a comprehensive analysis of worldwide Y-chromosome variation, it has been proposed that populations occupying the general area including Lake Baikal (eastward to the Trans-Baikal and southward into Northern Mongolia), the Lena River headwaters, the Angara and Yenisey River basins, the Altai Mountain foothills, and the region south of the Sayan Mountains (including Tuva and western Mongolia) was the source for dispersals of New World Y-chromosome founders (Karafet et al. 1999). It is obvious that we have now the genetic evidence that will allow closer determination of which Siberian population was the source of the population expansion leading to modern American Indians and will allow relation of the studies of migrations from Siberia to the Americas that are based on paternally inherited genetic systems with those based on maternally inherited ones.
Acknowledgments
The authors would like to thank the following individuals for providing samples for this study: Drs. E. Lotosh, F. Luzina, I. Dambueva, C. Dorju, V. Kakpakov, and O. Karamchakova. This work was supported by the Russian Fund for Basic Research (grant 99-06-80430), the State Program Frontiers in Genetics (grant 99-04-30), and by a grant from the Ludwik Rydygier Medical University in Bydgoszcz (BW 90/01).
Authors
MIROSLAVA V. DERENKO,1 TOMASZ GRZYBOWSKI,
2 BORIS A. MALYARCHUK,1
YAKUB CZARNY, 2 DANUTA
MIS´CICKA-S´LIWKA, 2 ILIA A. ZAKHAROV
3
1 Genetics Laboratory, Institute of Biological Problems of
the North, Magadan, Russia;
2 The Ludwik Rydygier Medical University in Bydgoszcz, Forensic Medicine
Institute, Bydgoszcz, Poland;
3 Animal Comparative Genetics Laboratory, Vavilov Institute of
General Genetics, Moscow
References
Alexeev VP, Gohman II (1984) Anthropology of Asiatic part of USSR. Nauka, Moscow
Anderson S, Bankier AT, Barrell BG, de Bruijn MHL, Coulson AR, Drouin J, Eperon IC, Nierlich DP,
Roe BA, Sanger F, Schreier PH, Smith AJM, Staden R, Young IG (1981) Sequence and organization of
the human mitochondrial genome. Nature 290:457-465
Bandelt HJ, Forster P, Sykes BC, Richards MB (1995) Mitochondrial portraits of human populations
using median networks. Genetics 141:743-753
Brown M.D., Hosseini S.H., Torroni A, Bandelt HJ, Allen JC, Schurr TG, Scozzari R, Cruciani F, Wallace
DC (1998)
mtDNA haplogroup X: an ancient link between Europe/ Western Asia and North America? Am J Hum Genet
63: 1852-1861
Calafell F, Underhill P, Tolun A, Angelicheva D, Kalaydjieva L (1996) From Asia to Europe:
mitochondrial DNA sequence variability in Bulgarians and Turks. Ann Hum Genet 60:35-49
Comas D, Calafell F, Mateu E, Perez-Lezaun A, Bosch E, Martinez-Arias R, Clarimon J, Facchini F,
Fiori G, Luiselli D, Pettener D, Bertranpetit J (1998) Trading genes along the Silk Road: mtDNA
sequences and the origin of Central Asian populations. Am J Hum Genet 63:1824-1838
Derenko MV, Denisova GA, Malyarchuk BA, Dambueva IK, Dorju CM, Stolpovski YM, Lotosh EA, Luzina
FA, Zakharov IA (2000a) Mitochondrial DNA variability in Turkic-speaking populations of the Altai
and Sayan region from South Siberia. Am J Hum Genet Suppl 67:A1161
Derenko MV, Malyarchuk BA, Dambueva IK, Shaikhaev GO, Dorju CM, Nimaev DD, Zakharov IA (2000b)
Mitochondrial DNA variation in two South Siberian aboriginal populations: implications for the
genetic history of North Asia. Hum Biol 72:945-973
Helgason A, Sigurdardo´ttir S, Gulcher JR, Ward R, Stefa´nsson K (2000) mtDNA and the origin of the
Icelanders: deciphering signals of recent population history. Am J Hum Genet 66:999-1016
Karafet TM, Zegura SL, Posukh O, Osipova L, Bergen A, Long J, Goldman D, Klitz W, Harihara S, de
Kniff P, Wiebe V, Griffiths RC, Templeton AR, Hammer MF (1999) Ancestral Asian source(s) of New
World Y-chromosome founder haplotypes. Am J Hum Genet 64:817-831
Lalueza C, Perez-Perez A, Prats E, Cornudella L, Turbon D (1997) Lack of founding Amerindian
mitochondrial DNA lineages in extinct Aborigines from Tierra del Fuego-Patagonia. Hum Mol Genet
6:41-46
Lell JT, Brown MD, Schurr TG, Sukernik RI, Starikovskaya YB, Torroni A, Moore LG, Troup GM, Wallace
DC (1997) Y chromosome polymorphisms in Native American and Siberian populations: identification
of Native American Y chromosome haplotypes. Hum Genet 100:536-543
Lutz S, Weisser HJ, Heizmann J, Pollak S (1998) Location and frequency of polymorphic positions in
the mtDNA control region of individuals from Germany. Int J Legal Med 111: 67-77
Luzina FA (1987) Hereditary polymorphism and genetic processes in indigenous populations of Altai
region. PhD thesis, Moscow State University, Moscow
Malyarchuk BA, Derenko MV (2000) Mitochondrial DNA variability in Eastern Slavs. In: Renfrew C,
Boyle K (eds) Archaeogenetics: DNA and the population prehistory of Europe. Cambridge, McDonald
Institute for Archaeological Research, pp 249-254
241
Orekhov V, Poltoraus A, Jivotovsky LA, Spitsyn V, Ivanov P, Yankovsky N (1999) Mitochondrial DNA
sequence diversity in Russians. FEBS Letters 445:197-201
Osipova LP, Kashinskaya YO, Posukh OL, Ivakin EA, Kryukov YA (1997) A genetic demographic study of
the South Altaian Population of the Mendur-Sokkon Village, Altai Republic. Genetika 33:1559-1564
(in Russian)
Parson W, Parsons TJ, Scheithauer R, Holland MM (1998) Population data for 101 Austrian Caucasian
mitochondrial DNA d-loop sequences: application of mtDNA sequence analysis to a forensic case. Int
J Legal Med 111:124-132
Piercy R, Sullivan KM, Benson N, Gill P (1993) The application of mitochondrial DNA typing to the
study of white Caucasian genetic identification. Int J Leg Med 106:85-90
Potapov LP (1969) Ethnical structure and origin of Altaians. Nauka, Leningrad
Rousselet F, Mangin P (1998) Mitochondrial DNA polymorphisms: a study of 50 French Caucasian
individuals and application to forensic casework. Int J Legal Med 111: 292-298
Santos FR, Pandya A, Tyler-Smith C, Pena SDJ, Schanfield M, Leonard WR, Osipova L, Crawford MH,
Mitchell RJ (1999) The Central Siberian origin for Native American Y chromosomes. Am J Hum Genet
64:619-628
Stone AC, Stoneking M (1998) mtDNA analysis of a prehistoric Oneota population: implications for
the peopling of the New World. Am J Hum Genet 62:1153-1170
Sukernik RI, Schurr TG, Starikovskaya YB, Wallace DC (1996) Mitochondrial DNA variation in native
Siberians, with special reference to the evolutionary history of American Indians: studies on
restriction endonuclease polymorphism. Genetika 32:432-439 (in Russian)
Torroni A, Huoponen K, Francalacci P, Petrozzi M, Morelli L, Scozzari R, Obinu D, Savontaus ML,
Wallace DC (1996) Classification of European mtDNAs from an analysis of three European populations.
Genetics 144:1835-1850
Underhill PA, Jin L, Zemans R, Oefner PJ, Cavalli-Sforza LL (1996) A pre-Columbian Y
chromosome specific transition and its implications for human evolutionary history. Proc Natl Acad
Sci USA 93:196-200
Wallace DC (1995) Mitochondrial DNA variation in human evolution, degenerative disease and aging.
Am J Hum Genet 57:201-223
Address for correspondence and reprints: Dr. Miroslava V. Derenko, Genetics Laboratory, Institute
of Biological Problems of the North, Portovaya Street 18, Magadan, 685000, Russia. E-mail:
ibpn (at) online.magadan.su
mderenko (at) mail.ru
T
T
P
.
Bibliography