Posting Foreword
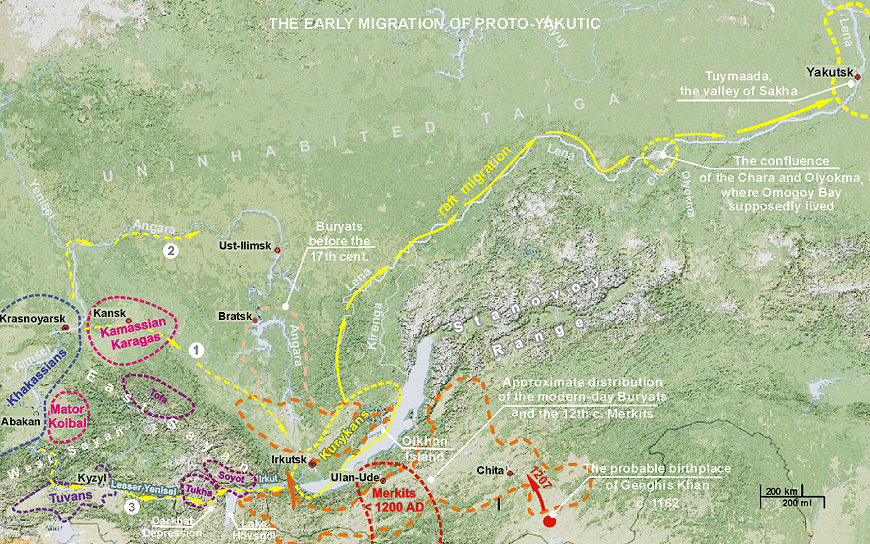
Of many Türkic migrations, the migration of Sakha people was established with a tip of a pen, and apparently, but not conclusively, we know both the departure and destination points. The following posting, preceded by genetic overwiew, aggregates findings from linguistical and historical disciplines.
Posting notes and explanations, added to the text of the authors and not noted specially, are shown in (blue italics) in parentheses and in blue boxes.
Origin of Sakha: Analysis of Y-chromosome Haplotypes
Molecular Biology, 2008, Volume 42, No 2, p. 226-237, © 2008
RESULTS AND DISCUSSION
Frequencies of Y chromosome haplogroup in Sakhas
Only some investigated biallelic marker loci on the results of genotyping of samples were informative for investigated Sakha samples, a total of only six haplogroups (Table 1). As expected, most common haplogroup in all the investigated samples was haplogroup N1c1 (formerly N3a) with almost 90% frequency in the total sample. It is the only haplogroup, identified in all three populations (3 villages). Areal of this haplogroup is the territory of Northern Eurasia, it is found in almost all modern Siberian ethnic groups. This haplogroup is not observed in the populations of Southeast Asia, the Americas, the Middle East: even in Mongolia and Central Asia it is only accidental.
V.A. Stepanov et al.
Origin of Sakha: Analysis of Y-chromosome Haplotypes
Molecular Biology, 2008, Volume 42, No 2, p. 226-237, © 2008
| Haplogroup | Frequency of occurrence,% (K) | |||
|---|---|---|---|---|
| All (K = 109) | Cheriktey (K = 46) | Byadi (K = 35) | Dyupsya (K = 28) | |
| С3хМ77 | 1.83 (2) | 2.17 (1) | - | 3.57 (1) |
| С3с | 1.83 (2) | - | - | 7.14 (2) |
| N* | 0.92 (1) | 2.17 (1) | - | - |
| N2 | 2.75 (3) | - | 8.57 (3) | - |
| N1c1 | 88.99 (97) | 89.13 (41) | 91.43 (32) | 85.72 (24) |
| R1a1 | 3.67 (4) | 6.53 (3) | - | 3.57 (1) |
| Н | 0.2071 ± ± 0.0518 | 0.2048 ± ± 0.0776 | 0.1613 ± ± 0.0787 | 0.2672 ± ± 0.1067 |
| H YDNA | 0.7911 ± ± 0.0422 | 0.8010 ± ± 0.0577 | 0.7311 ± ± 0.0818 | 0.8400 ± ± 0.0701 |
H - genetic diversity of haplogroups;
H YDNA - genetic diversity of microsatellite haplotypes.
The origin and distribution of this haplogroup is in circum-arctic zone of Eurasia. In Europe, haplogroup N1c1 with greatest frequency is found among Finns (60%), Saami (40-60%), Estonians (30%), and Finnish-and Turkic-speaking peoples of the Itil-Ural region (20-55%), Balts (30-40%), and East Slavs (5-15%). In southern, central and northern Europe N1 (formerly N3) occurs at a much lower freqiency.
In Siberia, haplogroup N1c1 comprises significant part in the male line of the Ural
peoples: Khanty, Mansi, Nenets (up to 40%). Significant proportion is found in
Buryats, and Chukchi (50%), Tuvans and Koryaks (up to 25%), Evens and Evenks
(15-40%). The highest frequency of haplogroup N1c1, after the Sakha, was recorded in
eastern Buryats (up to 85%) (own unpublished data). This line is found in almost all
modern Siberian ethnic groups, but not among indigenous populations of the New World.
Consequently, N1c1 reached the geographic source of migration to the Americas, the
Southern Siberia and Central Asia, later than American migrants headed for the Bering
Strait.
230
Each of the lines N*, N2, and C3c was detected only in one village - respectively in the village. Cheriktey, village Byadi, and village. Dyupsya.
According to the literature, the highest frequency of haplogroup N2 is observed in Nganasans (92%) and Nenetses (57%). In addition, from the Siberian ethnic groups a large proportion of N2 have Khanty (own unpublished data) Tuvas and Dolgans (12%). Among other ethnic groups the frequency of N2 does not exceed a few percentage points. In Europe, the range of N2 haplogroup is restricted to Itil-Ural region with about 10% of Y-chromosome at Mari, Chuvashes, Komi (13%) and Udmurts (29%).The haplogroup N* (N x M128, P43, TA1) is sporadically found in Siberia (mostly in the indigenous populations of Southern Siberia). The maximum frequency of this haplogroup is observed in populations of South-East Asia. Its appearance in the territory of Northern Eurasia may be connected with both N2 and N1c1, as well as with earlier migrants, which in our opinion is most likely. Apparently, N* line marks the migration route of Paleolithic migrants from Southeast Asia to the north, where in turn occurred mutations that define its derivatives, the sister lines N2 and N1c1.
| Population of Central China prior to the advent of Zhou Scythians (Xia period, ca
2300 BC) is identified as mostly Southeast Asian. All events of that period are described
as happening between the Jun (Jung) people, which apparently stands for Huns, and the
local defenseless agrarians of Southeast Asian origin. The morphed Southeast Asian-Zhou
compact was assailed from the east by ancestors of the modern Chinese (native Taiwan
type). The language of Yin is a controversial subject, ethnologically the Yin elite was
decidedly nomadic Türkic, the majority of population was Melanoid, some tribes were
Chinese. The Yin inscriptions are vaguely defined to be in a language of South Asian
type, but since the Yin script has developed into the Chinese script, in the Chinese
historiography the script is held as positively Proto-Chinese, and the language as
possibly Proto-Chinese, though all sources skillfully dance around the subject. There were two Shang Dynasties, one archaeologically described as Melanoid Qiang-Shang and the other as Polynesian Yin-Shang. The first Shang Dynasty was alternatively founded by Melanoids belonging to the Yueh tribe called Qiang 羌 from the Qiangfeng 羌方, a country to the west of Yin-Shang, Shensi and Yunnan. The Melanesian and Polynesian populations lived in intimate contact for millennia and probably heavily exchanged genes. The Polynesians probably originated in East Asia, not Southeast Asia. Genetically, Taiwan probably belongs to the early Polynesians who settled Taiwan before they expanded into outer Oceania. By the 1000 BC the Hau/Han tribes came down from the mountains and began enslaving Yueh and Li Min people, killed off as many Yueh, Qiang and Li Min tribes as they could, often used Qiang as sacrifice victims, and pushed the classical Mongoloids southward into Yunnan and eventually Southeast Asia. The Han are ancestors of the Chinese Han people. This movement of Han and classical Mongoloid people southward forced the Qiang, Li Min and other Melanesian tribes onto the Pacific coast. The Chinese and Classical Mongoloid people share few if any genes with the Australians. The Classical Mongoloids share genes mainly with the coastal Melanesians who are of African origin, but few genes with the Chinese of East Asia. Speculatively, the push of Hau/Han tribes could be an impetus for Southeast Asians exodus not only southward, but also northward. |
Haplogroup C3 x M77 occurs with the highest incidence in the territory of Central Asia and Eastern Siberia, where its carriers diffused the from Southeast Asia.
The line C3c is mainly spread in the Central and South Central Asia and Siberia populations, but not in Southeast Asia and America. Such phylogeography suggests a later date, relative to C3 x M77, of its spread to the continent and its probable origin in the Central Asian region.
The final component of the Sakha Y-chromosome gene pool is haplogroup R1a1 (3.67% of total sample). This line is found with varying frequency in most South Siberian populations and is associated with Caucasoid component in their gene pool. Distribution of carriers R1a1 eastward went through the Eurasian steppe belt, up to the northern Mongolia, where were found singular chromosomes belonging to this haplogroup. Analysis of the literature on YSTR-haplotypes shows that majority of R1a1 chromosomes belonging to Central Asia and southern Siberia ethnic groups with that haplogroup are a single genetic array, in stark contrast in composition and frequencies of haplotypes to the European chromosomes. This indicates a relatively early separation of the carriers R1a1 into "Europeans" and "Asians". We surmise that this haplogroup (although present in this territory previously) was spread with migrations of the ancient Türks in the Southern Siberia as well as outside of it. In the future, likely will be found new informative biallelic markers, which combined with genotyping of a large quantity of YSTR markers, will allow to determine within R1a1 a specific range of lines associated with the separation of the carriers of the Turkic languages. The low frequency of this haplogroup in Sakha is quite interesting, because in the gene pools of all the Turkic-speaking ethnic groups of Southern Siberia it is a significant part. This difference in composition between the Sakha haplogroups and their more southern neighbors indicates the formation of Y-chromosomal part of their gene pools from different by origincomponents, and possible absence of close inter-ethnic contacts between them. Obviously, the Turkic language was acquired by the ancestors of modern Sakha from the outside, perhaps as a result of cultural dominance by the migrant Turkic elite.
All investigated Sakha samples are characterized by very low value of genetic diversity, calculated on the basis of haplogroup frequencies (Table 1). Genetic diversity indicators for other Siberian ethnic groups are much higher. Given that Sakha are one of most numerous indigenous peoples of Siberia, and that they are presented in our study by three population samples, and the similar results obtained by other authors, the scenario with gradual reduction in genetic diversity due to genetic drift seems unlikely. Rather, it was a founder effect. This premise is supported by mtDNA data: its distinctive feature is the predominance of Sakha mtDNA types belonging to haplogroups C and D.
Genetic differentiation of populations
...Thus, the analysis of microsatellite haplotypes of Y chromosome demonstrates more clearly the differentiation of the Sakha sample, both in accurate testing of population differentiation, and by the molecular variance (AMOVA). This is due to the predominance in the Sakha gene pool of the line N1c1. ...population samples are distinct in larger value of diversity indicators and greater degree of differentiation.
Phylogenetic analysis of haplogroup N1c1a in Sakhas
Case with a Y-chromosome line in such significant part of the gene pool in individual
population or ethnic group is generally quite rare. In no modern population was the haplogroup N1c1
found with greater frequency than among Sakha. ...Phylogenetic analysis of N1c1 haplogroup with
YSTR markers allows to establish detailed structure by
constructing a haplotype tree using method of median networks. ... a dataset for frequencies and
structure of haplotypes
within the Y-chromosome N1c1 haplogroup in the populations of the
world: 172 Sakha, 24 Evenks, 5 Mongols, 5 Turks, 72 Lithuanians, 17 Latvians, 30 Estonians, and... 5
Southern Altaians (Tuvinians), 14 Buryats from Agin
Autonomous District, 7 Buryats from Buryatia Republic, 19 coastal Chukchi, 11 Evenki from Chita
Province, 6 Khanty, 8 , 9 Ukrainians, 7 Belarusians, 26 Russians, 11 Siberian Tatars and
51 Tuvans. ... for seven
markers in the present study.
...accepted N1c1 haplogroup founder haplotype
14-11-16-23-11-14-14 (Tuvinians,
Altaians, Chukchi, Koryaks, Buryats,
Siberian Tatars)
...accepted Sakha founder haplotype
14-11-18-23-11-16-14.
Almost all Sakha and Evenki haplotypes are derivatives and not directly related to the network common founder haplotype. ... Altogether at Sakha were found forty haplotypes of N1c1 haplogroup. Nine out of thirteen Evenki haplotypes coincide with the Sakha haplotypes, differing only by relative frequency, the remaining four are separated from them by a various number of mutations.
To summarize, Evenks and Sakhas have very specific range of N1c1 haplotypes, largely overlapping, indicating that a part of gene pools of these two ethnic groups formed from a common base. Moreover, the nine Sakha haplotypes coincide with haplotypes found in other ethnic groups: two each at Tuvinians and Buryats and one each at Khanty, Koryaks, Mongols, Chukchi, and Kirgiz. Eight of these haplotypes are separated from the entire network common founder haplotype by no more than two steps, and therefore belong to a different genetic pattern. The overall percentage of Sakha samples with these haplotypes is just over 3% of all haplotypes belonging to N1c1.
The majority of the remaining haplotypes, with exception of only five, are clustered around ancestral Sakha haplotypes at a distance of two mutational steps, and there are no "empty" nodes...This obviously indicates that the demographic history of Sakha encountered a sharp reduction in the effective number of ancestral populations, perhaps initiated by a small group of founding migrants, whose gene pool had little diversity and a shift of the haplotype spectrum relative to the general population, apparently similar to Buryat and Tuva.
Buryats1 - sample from Buryatia Republic; Buryats2 - sample from Agin Autonomous Region of Chita
Province;
Evenki1 - sample from Chita Province; Sakha1 - sample from Ust-Aldan ulus of
Sakha Republic;
Evenki2 and Sakha2 - sample studied in Pakendorf at al.
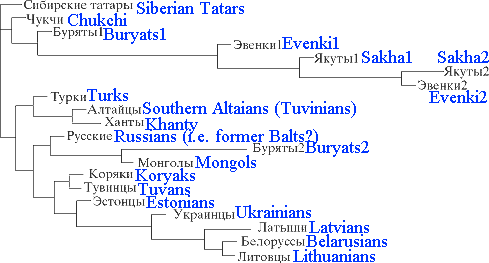
| Somehow this dendrogram does not look right, even allowing that under politonym Russians are hidden Balts. To allow that closest to the Byryats and Mongols are Russians (Russian Balts), and at the same time they are twwo steps away from the Turks (of Anatolia?) seems to be unreal. |
235
Ethnogenetic reconstruction
...Most feasible in our opinion is the following scenario. The Sakha (more accrately the Turkic tribes participating in forming the Sakha ethnic group) and Evenki (Tungus tribes) got their N1c1 haplogroup relatively recently from an external source, with practically already established haplotype variety observed at present time. So, in the territory of modern Sakha, and perhaps beyond it, lived very closely related populations whose gene pool had a very high frequency of N1c1 haplogroup with a very unique range of haplotypes.
Determine the range limits of these populations more accurately seems to be impossible, but the territory of E.Baikal should be a part of it, because our Evenki sample is formed in the Chita Province. Separation of these tribes from ancestral community of all ethnic Siberian groups N1c1 according to our calculations happened about five thousand years ago. These populations were assimilated after migration of Tungusic tribes, which settled in Sakha area at the end of the 1st - the beginning of the 2nd mill. AD. (which led to the formation of modern Evens and Evenks), and later by Türkic-speaking migrants who probably have assimilated them (Tungusic tribes) and Evenks (which led to formation of the nucleus of Sakha ethnic group).
| That scenario is consensual, except for the dating, see reference above, which should not alter the linguistic picture. The difference between 700 ya and 1300 ya is quite significant, and may point that the constants used for dating require calibration; otherwise the flow of historical events may have happened differently. |
The shares of the ancient Paleolithic tribes contribution to the Y-chromosome gene pool component of the Evenks and Sakhas was not identical: at Evenks it is significant, at Sakhas it is overwhelming (That essentially means that descendents of ancient Paleolithic tribes, read Yukagirs, were inseminating the Türkic refugee females and their descendents, pointing to the predominantly female Türkic exiles either 700 or 11300 years ago). The relative stability of Evenks' demography from the time they received that component provided preservation of the frequencies for individual haplotypes, probably close to the initial frequency distribution. A significant increase in the Sakha numbers in the last few centuries has led to accidental sharp increase in frequencies of some haplotypes, predominantly of probable founder haplotype. A similar picture can be seen in the Sakha mitochondrial gene pool: predominance in frequency of some haplogroup C haplotypes, and very high frequency of one haplotype in the haplogroup D.
Apparently, Yukagirs, or more accurately Proto-Yukagirs, and closely related to them Paleo-Asiatic tribes in the Sakha territory unknown to us by their ethnonym are candidates for the role of the donor population. A few questions remain unclear.
1) Did the highly specific range of haplotypes of the N1c1 haplogroup observed at the present Sakha (or some of its part) belong to the gene pool of the Türkic-speaking migrants before their arrival in the Sakha territory, or they received it in full only later.
2) Is such structure of haplogroup N1c1 typical for other Evenk and Even populations throughout their range, and for populations of other ethnic groups, particularly, for western Buryats and reindeer Chukchi.
3) To what degree that structure was typical for Yukagirs.
...The second
scenario (lack of cluster-specific haplotypes among Türks before their coming to the
Sakha territory) has higher probability because South Siberian Türkic ethnic groups have
quite different range of haplogroup N1c1 haplotypes. ...To resolve these issues, additional samples of Evenks, Evens, Buryats, Chukchi and deer
Sakha should be examined, and other additional biallelic markers should be used, allowing to establish
detailed molecular phylogeny of N1c1 haplogroup in Siberia....
236
...It should be emphasized that vision of the Sakha as "Turkified" Yukaghirs, who formed the locally-based overwhelming majority of the entire gene pool is fundamentally wrong. Such interpretation would be overly simplistic. The data on the mtDNA nuclear markers of the A1u-repeats, and biochemical markers confirm significant contribution to the Sakha gene pool of by the components of southern origin that converge Sakha with the South Siberian and Central Asian populations. Finally, the significant anthropological differences between Sakha on the one hand, and Yukagir and Evenks on the other hand, perhaps most graphically demonstrate the importance and significant contribution of the migrant component to the formation of the Sakha ethnic group.